
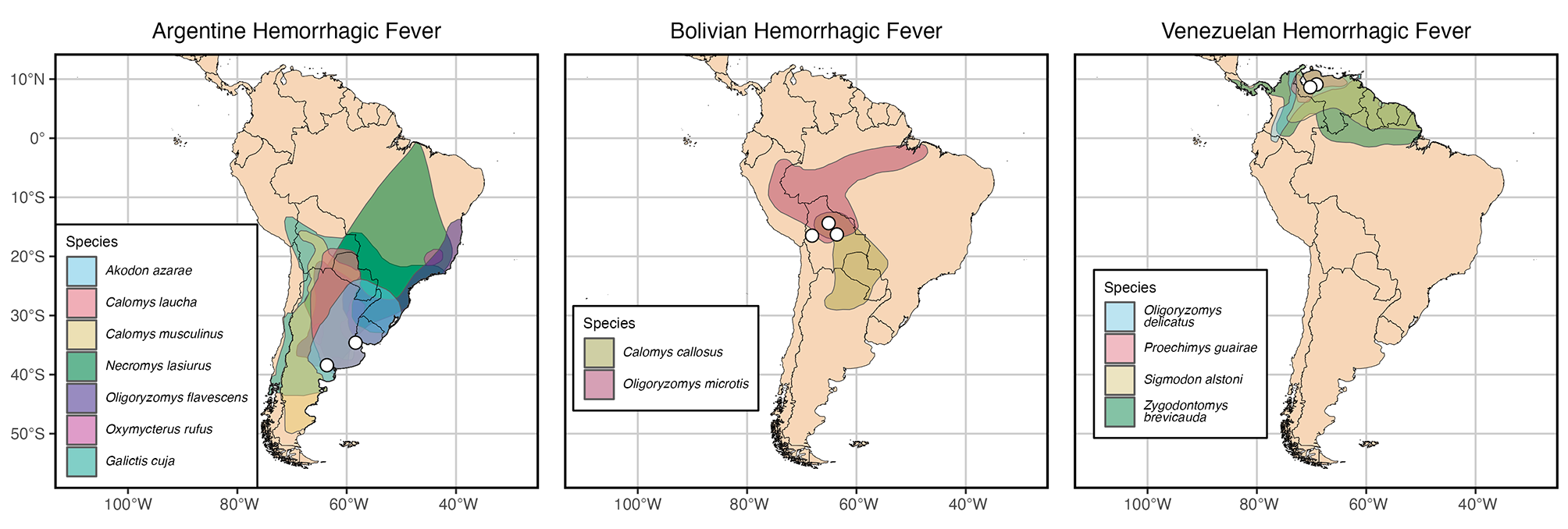
記住我

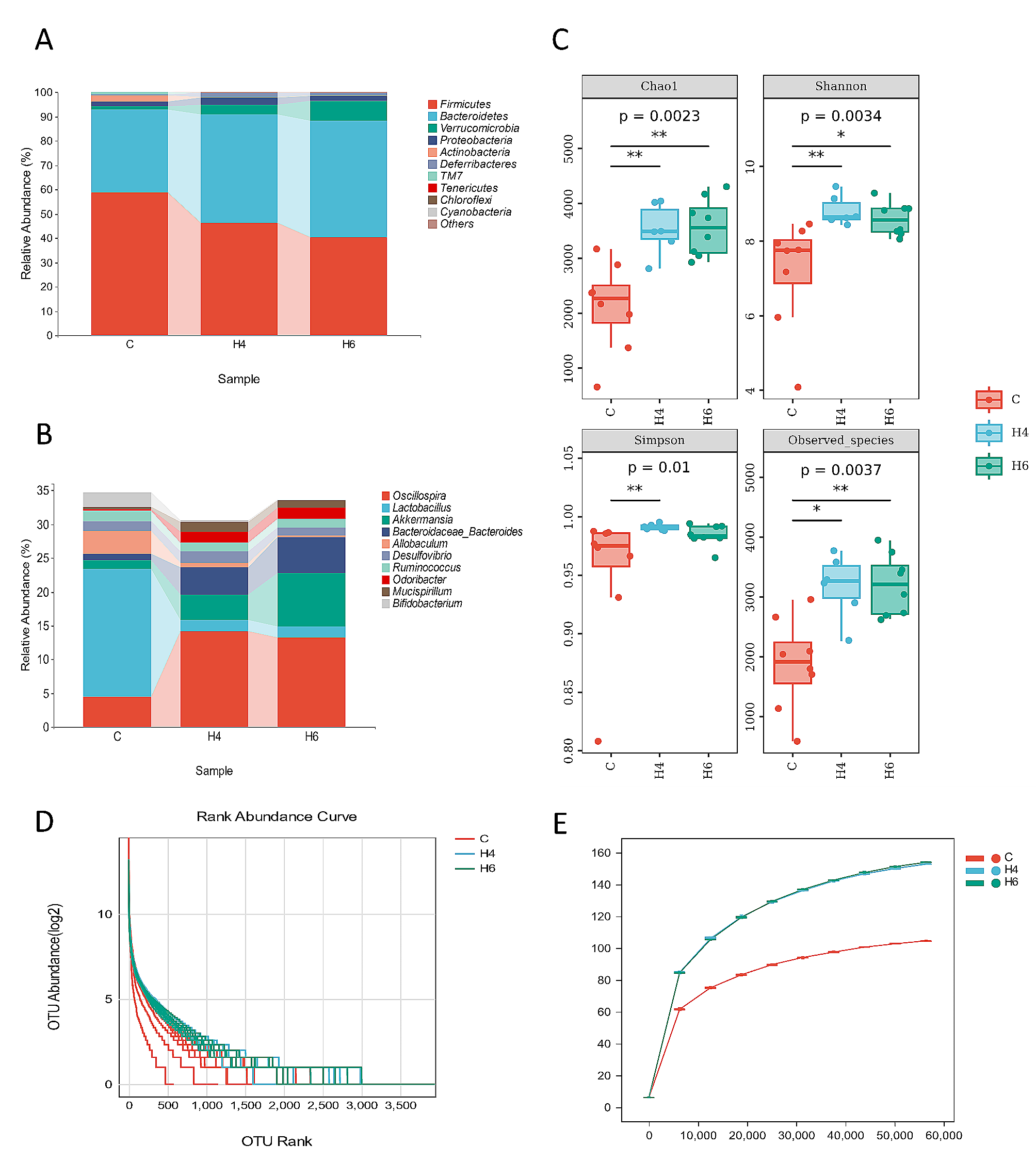
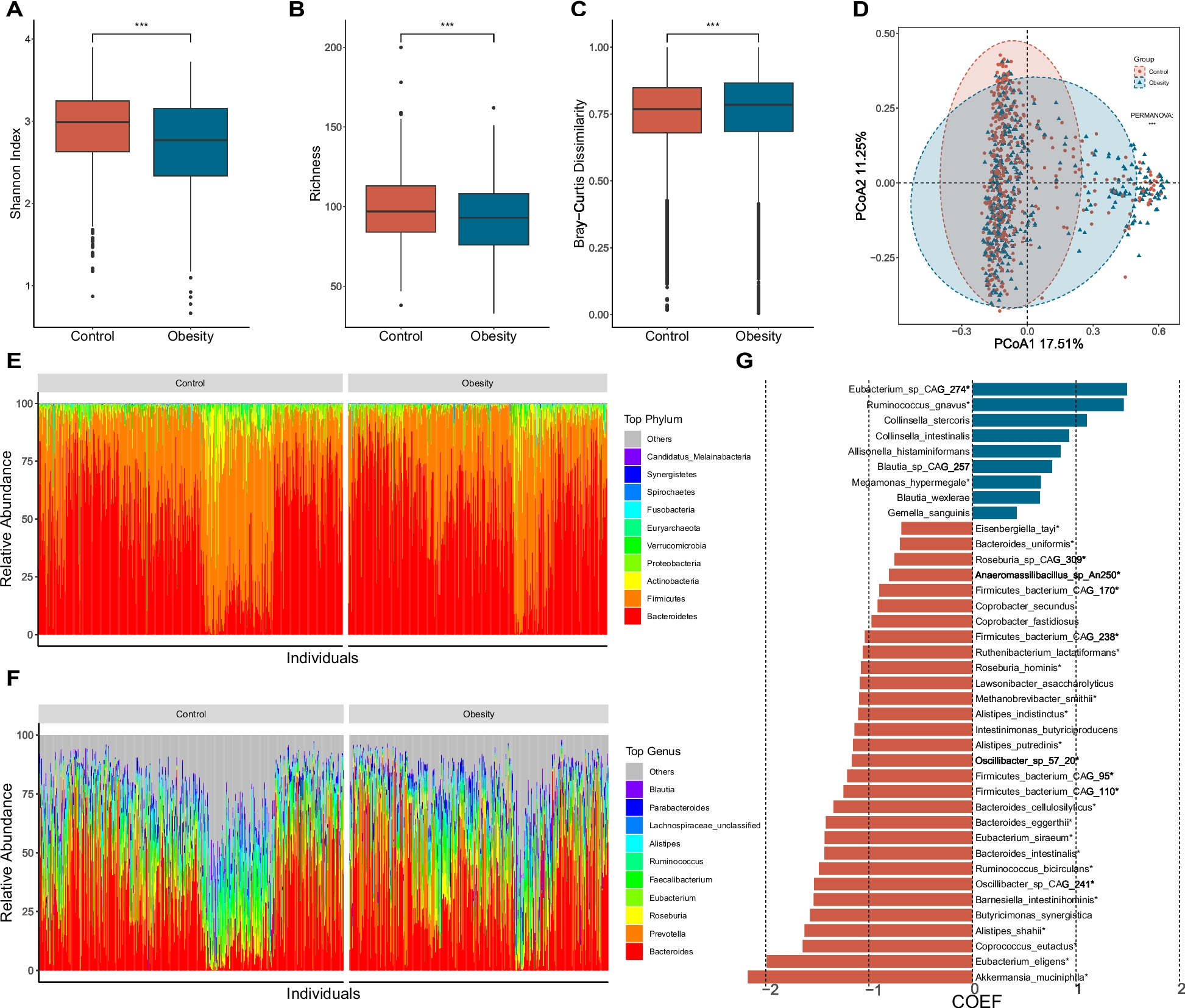
Under the screening condition of FDR < 0.05 and |log2FC|> 2, a total of 756 Unigenes were selected. The gene sequences from assembled transcriptome data against the NR database resulted in the identification of β-glucosidase genes, bgl3HB, under the stringent criteria of at least 50% identity and 90% subject coverage. This putative β-glucosidase gene, in turn, did not reveal similarity with any of the sequences available in NCBI non-redundant nucleotide database on the default BLASTn parameters. This establishes the novelty of the β-glucosidase gene characterized in this study. The molecular weight and theoretical isoelectric point of Bgl3HB was predicted to be 86.66 kDa and 5.35, respectively, by the ExPASy website (https://web.expasy.org/compute_pi/). The SignalP5.0 and TMHMM2.0 analyses showed that the gene sequence of Bgl3HB had no signal peptide and belonged to an exocrine protein.
The domains of Bgl3HB were analyzed with the SMART website (SMART: Main page (embl-heidelberg.de)). As shown in Fig. 1, Bgl3HB contains a Pfam Glyco-hydro-3 domain, a Pfam Glyco-hydro-3-C domain and a Fn3_like domain. Glyco_hydro_3 and Glyco_hydro_3_C domains belong to the Glycoside hydrolase family 3 (GH3) and participate in catalysis. GH3 comprises enzymes with a number of known activities; β-glucosidase ( EC 3.2.1.21); β-xylosidase (EC 3.2.1.37); N-acetyl β-glucosaminidase (EC 3.2.1.52); glucan β-1,3-glucosidase (EC 3.2.1.58); cellodextrinase (EC 3.2.1.74) and exo-1,3–1,4-glucanase (EC 3.2.1). These enzymes are two-domain globular proteins that are N-glycosylated at three sites. This suggests that Bgl3HB belongs to the GH3 family, with a Fn3-like domain in the C-terminal. The specific function of Fn3-like domain is unknown, which may be related to the thermal stability of the protein.
Fig. 1
Domain analysis of Bgl3HB
Expression and purification of Bgl3HBRecombinant Bgl3HB was successfully produced in K. phaffii X33 cultured cells in BMMY medium. Figure 2 showed the changes of the enzyme specific activity of the supernatant of the recombinant strain and the control strain under the induction of 1% methanol for 7 days. The mass spectrometry identification results are shown in Fig. 3.
Fig. 2
Changes in specific activity of Bgl3HB in fermentation supernatant for 7 days
Fig. 3
From the position of the band, it can be judged that Bgl3HB is correctly expressed (Fig. 4), given the similarity to the expected protein molecular weight.
Fig. 4
The molecular weight and purity of Bgl3HB were confirmed by SDS-PAGE on an 8% SDS gel. Lane 1: PageRuler Prestained Protein Ladder (Thermo Scientific, USA); and Lane 2: purified Bgl3HB stained by Coomassie Brilliant Blue BL605A
Biochemical characteristics of recombinant Bgl3HBThe enzymatic properties of purified recombinant Bgl3HB produced in K. phaffii X33 were determined using pNPG as the substrate.
Purified Bgl3HB was active over a wide range of acidic pH, exhibiting optimal activity at pH 4.0 and retaining > 76% activity at pH 3.0–5.0 (Fig. 5a). The pH stability was investigated by measuring the residual activity after incubation at 4 °C, at pH values ranging from 3.0 to 5.0. As shown in Fig. 5b, Bgl3HB was stable at pH 4; it retained more than 73% of its residual activity after 240 min of incubation in pH buffer at 4 °C. Most fungal β-glucosidase show pH optima ranging from 4.0 to 6.5 and are usually stable over a wide pH range [20, 21]. This optimum pH and stability indicate that Bgl3HB have potential commercial value, because stability for a long time under extreme pH conditions is one of the most desired industrial properties of β-glucosidase. β-glucosidase that can tolerate low pH and remain relatively stable are very advantageous [22] in the use of pretreated lignocellulose slurry, which is usually acidic. In practical production applications, such as in the wine-making process, acid-resistant β-glucosidase plays a key role in the enzymatic release of aromatic compounds from glycoside precursors present in fruit juices and wines [23]. Acid-tolerant β-glucosidase is very important in the food and beverage industry and in the production of fuel ethanol from cellulosic materials [24].
Fig. 5
Enzymatic properties of the purified recombinant Bgl3HB produced in K. phaffii using pNPG as the substrate. a Effect of pH on enzyme activities. b pH stability of Bgl3HB after 240 min incubation at 4 °C. c Effect of temperature on enzyme activities. d Thermostability of Bgl3HB at pH 4.0 and different temperatures between 50 °C and 70 °C up to 240 min. Each value in the panel represents the mean ± SD (n = 3)
The optimum temperature of the hydrolysis reaction was found to be 50 °C (Fig. 5c). When Bgl3HB activity was assayed at pH 4.0, it exhibited maximum activity at 50 °C and retained 90% activity even at 60 °C (Fig. 5c). Bgl3HB was also highly stable at 50 °C, retaining > 65% activity after 240 min (Fig. 5d). Previous results of Aspergillus fumigatus Z5 [25], Trichoderma oranges [26] and Trichoderma mange [27] showed that thermal decomposition is usually used to promote the degradation of lignocellulose biomass, so heat resistance is an ideal property of β-glucosidase. And thermostable β-glucosidases offer several advantages in industrial applications, such as promoting faster reactions, high solubility of the substrate and a lower risk of contamination, which has led to an increased desire for a new type of β-glucosidase with high reaction temperature and thermal stability [28]. In this study, Bgl3HB can even maintain more than 50% enzyme activity after incubation at 60 ℃ for 120 min. Thermostable β-glucosidase not only plays an extremely important role in the high temperature growth of T. harzianum in its native mangrove wetland, but also thermostable β-glucosidase in industrial processes such as biorefining. If the substrate needs to be heat treated, thermophilic enzymes can increase the reaction rate, shorten the hydrolysis time, and have excellent specific activity of reducing the number of enzymes. Higher temperature stability and better pH flexibility generally related to process configuration [29, 30]. Because most commercial cellulases currently on the market do not have good thermal stability, and β-glucosidase is the rate-limiting enzyme in the process of cellulose hydrolysis, the demand for thermostable β-glucosidase is significant. First, the temperature of enzymatic hydrolysis of plant biomass is 30–50 ℃, so a thermally stable β-glucosidase is favorable in this enzymatic reaction. Second, enzymatic hydrolysis at higher temperatures can effectively avoid contamination of the reaction with undesirable bacteria. Therefore, the β-glucosidase produced by T. harzianum from Hainan mangrove in our laboratory has a wide range of commercial value and application prospects.
Effect of metal ions on enzymatic activityBgl3HB was highly resistant to all tested 1 mM and 5 mM metal ions (Fig. 6a), and no similar reports had been found. In particular, Ca2+ and Mn2+ extremely enhanced Bgl3HB activity by 30.96% and 29.98%, respectively. Collectively, these results indicate that most metal ions can promote the activity of Bgl3HB, which meets the needs of industrial production. In the process of cellulose saccharification, metal ions often inhibit the activity of glucosidase, so metal ions tolerance is a desirable feature for enzymatic bioprocesses [31].
Fig. 6
Effect of metals and NaCl on Bgl3HB enzyme activity with p-nitrophenyl-β-D-glucopyranoside (pNPG) as substrate. The values represent the mean ± SD (n = 3). a The reactions were performed by incubating purified enzyme with 1 mM and 5 mM of various metal ions (Cu2+, Ca2+, Ni2+, Mg2+, K+, Al3+, Mn2+, Zn+, Fe3+, Co2+) at 50 °C for 10 min; CT means the contrast group. b Enzyme was incubated in 0–5 M NaCl at 50 °C for 10 min and determined the specific activity of the enzyme. The highest enzyme specific activity was taken as equivalent to 100% specific activity
Effect of NaCl on enzyme activityThe effects of salt on the activity of Bgl3HB were determined at the concentrations of 0–5 mM (Fig. 6b). Notably, at higher concentrations of salt there was a stronger promotion of enzyme activity. To evaluate the activation level of Bgl3HB by NaCl compared to other β-glucosidases, we provide a summary of reported β-glucosidases that can be activated by NaCl in Table 1. Bgl3HB is the β-glucosidase exhibiting the highest NaCl activation concentration, and it is the first halophilic β-glucosidase from T. harzianum to be reported. This may be because mangroves are marine ecosystems, with high salinity and soil osmotic pressure, leading to salt stress tolerance of most microorganisms [17]. β-glucosidase isolated from mangrove soil has previously been shown to be salt tolerant, and enzyme activity can even be enhanced by salt concentrations. Most of the β-glucosidases isolated from oceanic microbes show similar properties. It has been reported that a cellulase produced by marine Aspergillus niger ZJUBE-1 isolated from the sludge of the East China Sea can retain nearly 80% of the enzyme activity in the presence of 4.5 M NaCl [32]. The β-glucosidase produced by Streptomyces from deep-sea sediments can stably exist in 1 M NaCl for 48 h [17]. In this study, Bgl3HB derived from mangroves was shown to tolerate NaCl up to 5 M (higher than seawater salinity), and its activity increased by 2 times. Microbes living in mangroves often have to cope with extreme salinity gradients caused by seasonal rainfall, tidal conditions and freshwater outflows [33]. In addition, the distribution of species is regulated by high, medium and low salinity regions, which can explain the tolerance of Bgl3HB to NaCl.
Table 1 Comparison of activity toward pNPG by β-glucosidases activated by NaClSubstrate specificity assays and kinetic parameters of Bgl3HBTo determine substrate specificity, the hydrolytic activity of purified Bgl3HB was measured using various substrates. We found that Bgl3HB showed extensive hydrolytic activity against many substrates. It had high specific activity of 58.34 U/mg on daidzein and 129.79 U/mg on laminarin. When using disaccharides of different linkages as the substrate, the enzymes showed different levels of preference, in the order of Gentiobiose (β-1,6 linkage) > Sophorose (β-1,2 linkage) > Laminaribiose (β-1,3 linkage) > Cellobiose (β-1,4 linkage) > Cellopentaose (β-1,4 linkage) > Cellotetraose (β-1,4 linkage) > Cellotriose (β-1,4 linkage) (Table 2). And for various aryl-glycoside substrates, Bgl3HB was active with daidzein and laminarin as preferred substrates. Surprisingly, it also showed a remarkable ability to hydrolyze laminarin, a β-1,3-glucan present in algae, which makes Bgl3HB a good candidate for using in the production of 3G biofuels from algal biomass [37]. Recently, numerous algae species have been studied for this objective. For example, green algae, including Spirogyra sp. and Chlorococcum sp., accumulate high levels of polysaccharides like starch, which could be fermented to bioethanol [38]. Méndez-Líter found a β-glucosidase from the ascomycete fungus Talaromyces amestolkiae, which had a higher hydrolysis rate than a commercial laminarinase for laminarin [37].
Table 2 Substrate specificity of purified recombinant β-glucosidase Bgl3HBThe kinetics of Bgl3HB on daidzein, laminarin and pNPG are shown in Table 3. Bgl3HB exhibited much higher substrate affinity and catalytic efficiency on laminarin than those of other Aryl-glycosides. It had the lowest KM value, which means that Bgl3HB had high affinity for laminarin. The versatility of some β-glucosidases to hydrolyze short oligosaccharides with β-1,2, β-1,3, β-1,4, or β-1,6 linkages has been widely reported [39]. However, the ability of Bgl3HB to degrade a polysaccharide like laminarin is truly exceptional, which makes Bgl3HB have great advantages in the production of biofuel with algae biomass as raw material. Recently, 3G biofuels derived from algae biomass have received considerable attention for their advantages as biodiesel, bioethanol, biohydrogen, and biomethane [40]. This catalytic property of Bgl3HB could be of great interest for depolymerization of glucans for 3G bioethanol production.
Table 3 Kinetic parameters of Bgl3HB with different substratesEnzymatic saccharification of cellulose materialsIn order to evaluate the application value of Bgl3HB under practical conditions, the application potential of Bgl3HB in saccharification of cellulose raw materials was compared with that of commercial Novozyme 188 at pH 4.0 and 50 ℃. As shown in Fig. 7, bagasse was used as raw material, in the blank control group, commercial Celluclast 1.5L (5 FPU (Determined by pre-experiment)/g dry material) was cultured at pH 4.0, 50 °C for 96 h, reducing sugars from bagasse released 296.4 μmol, of which glucose accounted for 42.9 μmol (the glucose conversion rate of 14.5%). When β-glucosidase was added at 12 BGU (Determined by pre-experiment)/g dry material, Bgl3HB and commercial Novozyme 188 showed different synergistic effects in promoting saccharification. The synergistic action of Celluclast 1.5L and commercial Novozyme 188 released 353.8 μmol of reducing sugar and 61.5 μmol of glucose (glucose conversion rate 17.4%). Bgl3HB in combination with Celluclast 1.5L only released 344.0 μmol of reducing sugars and 58.1 μmol of glucose (the glucose conversion rate of 16.9%). However, Celluclast 1.5L and commercial Novozyme 188 with additional 5 mM NaCl improved performance in synergistic enzymatic saccharification. The yields of reducing sugars and fermentable glucose were 380.84 μmol and 67.60 μmol (the glucose conversion rate of 17.8%), which is higher than that of commercial Novozyme 188.
Fig. 7
Enzymatic saccharification of Bgl3HB (12 BGU/g dry material) in combination with commercial cellulase (5 FPU/g dry material). The pretreated bagasse was used as the substrate. a The reducing sugar released by enzyme(s). b The glucose released by enzyme(s)
Notably, although the bagasse conversion rate of Bgl3HB is lower than that of commercial Novozyme 188, the glucose conversion rate is higher in Bgl3HB than in commercial Novozyme 188. However, the bagasse conversion rate of Bgl3HB with an additional 5 M NaCl was higher than that of commercial Novozyme 188, and the glucose conversion rate was also higher than that of commercial Novozyme 188. This agrees with other results in our study, suggesting that NaCl can increase Bgl3HB activity, and may help improve the hydrolysis capacity of Bgl3HB in the saccharification process. Such an effect may be because salt is conducive to the stability of the enzyme [16]. Qu et al. found that 3 M NaCl could improve the conversion rate of soybean isoflavones [34], which is similar to the findings of this paper. This important interaction would allow the hydrolysis of commercial cellulase with a smaller quantity of enzyme, which would reduce the industrial application cost.
ConclusionsHere, a β-glucosidase gene derived from T. harzianum HTASA was cloned and heterologously expressed in K. phaffii X33. To our knowledge, this is the first report of a β-glucosidase identified from T. harzianum HTASA. A detailed enzymatic characterization of Bgl3HB was then performed. Bgl3HB was found to be heat-resisted and salt-tolerant, and could hydrolyze efficiently laminarin, which indicated that Bgl3HB had important cellulosic biomass degradation. Collectively, Bgl3HB is a salt-enhanced β-glucosidase with broad pH stability, high thermostability, uninhibited properties by most metal ions, and broad substrate specificity, which makes it has potential applications in different biotechnological applications, including bagasse hydrolysis and bioethanol production, and provides a useful material for further research.
留言 (0)