
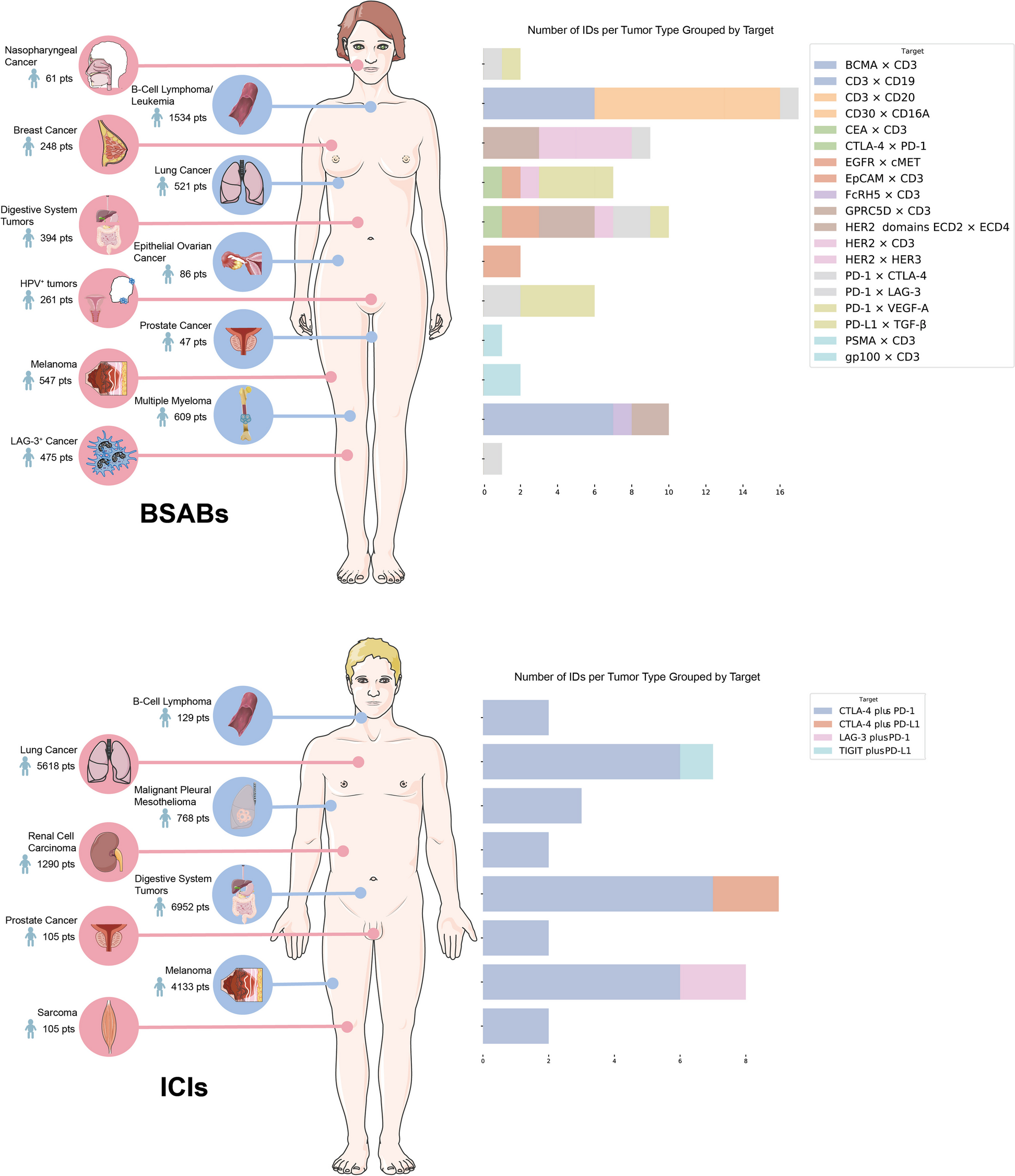
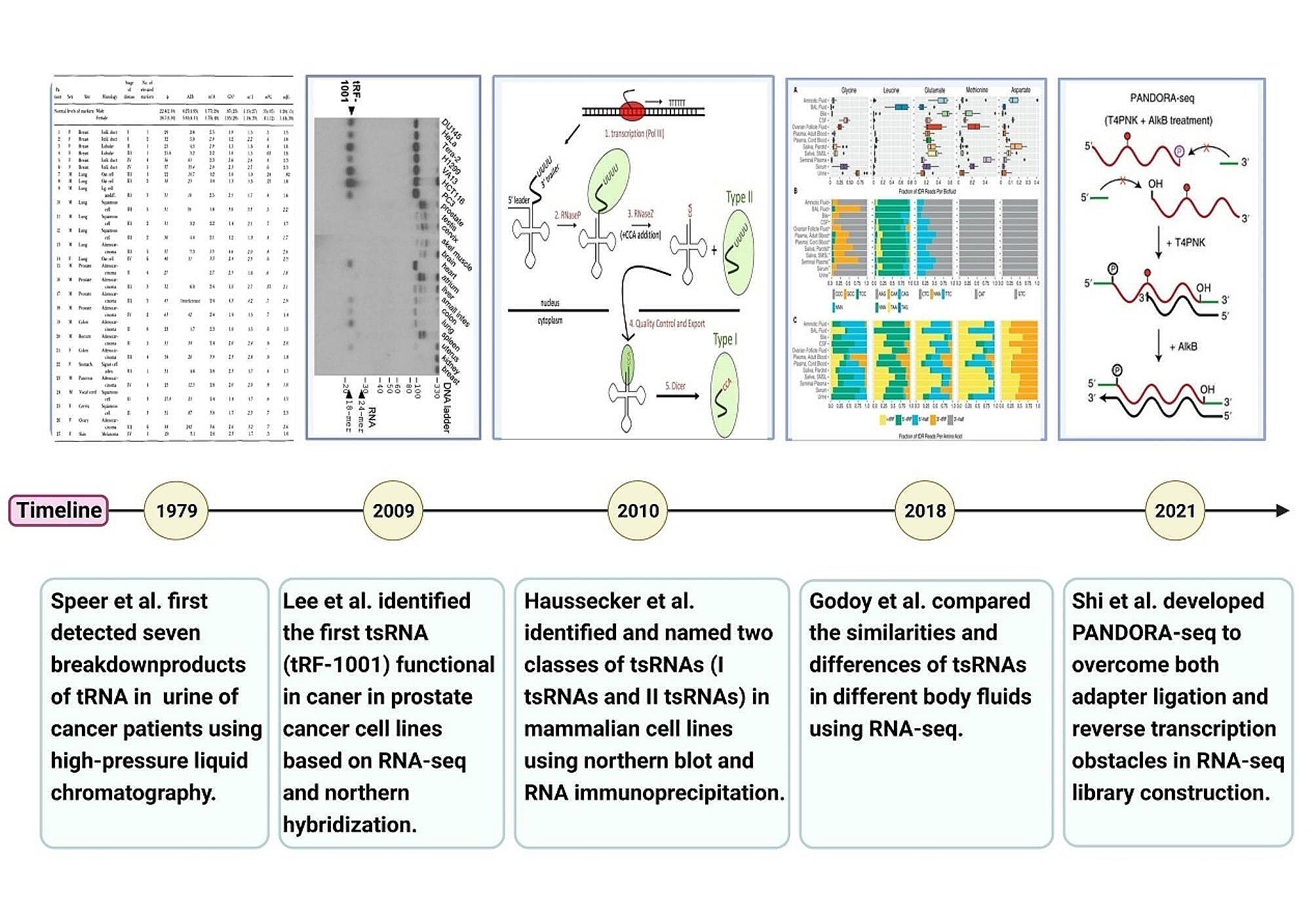
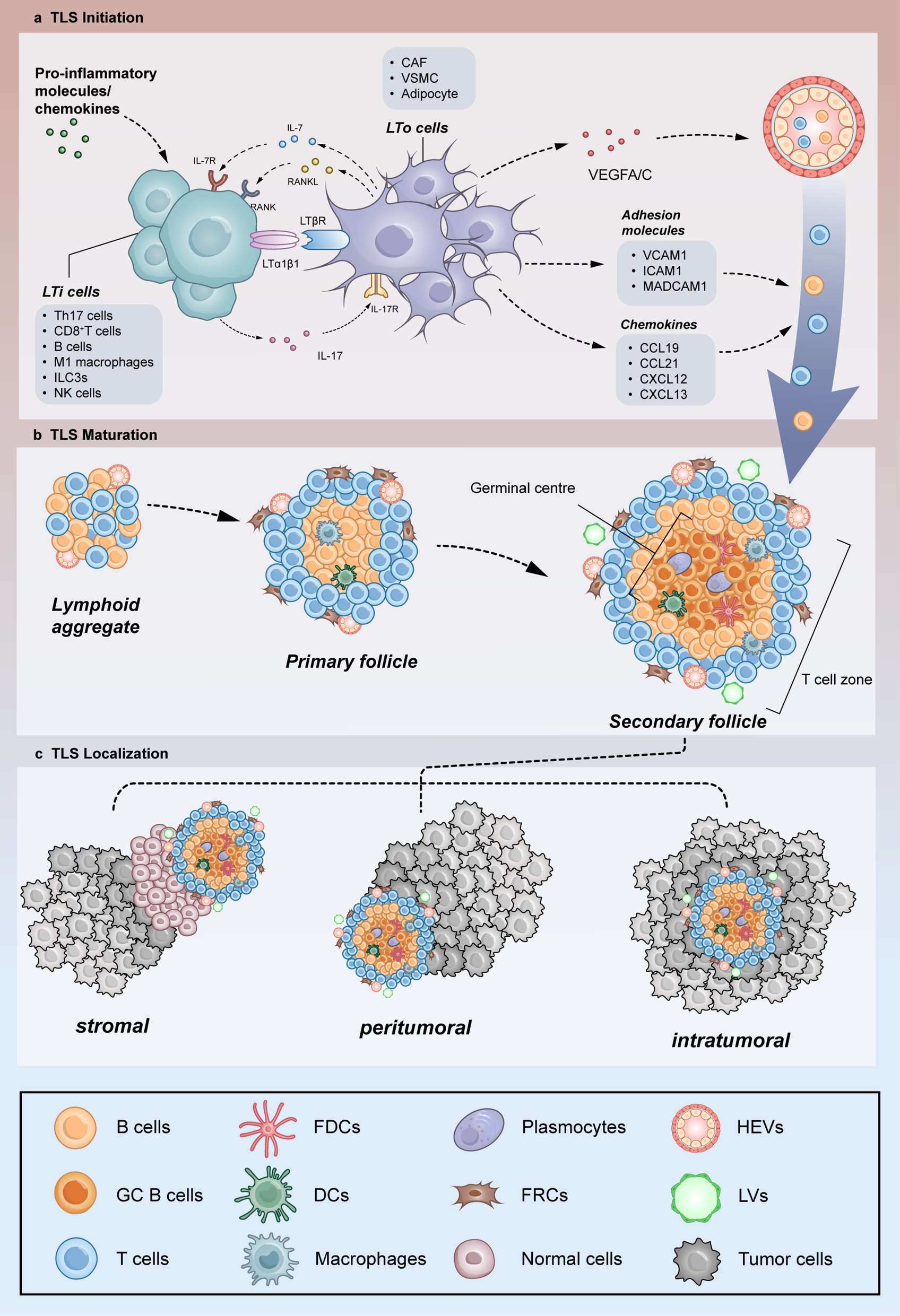
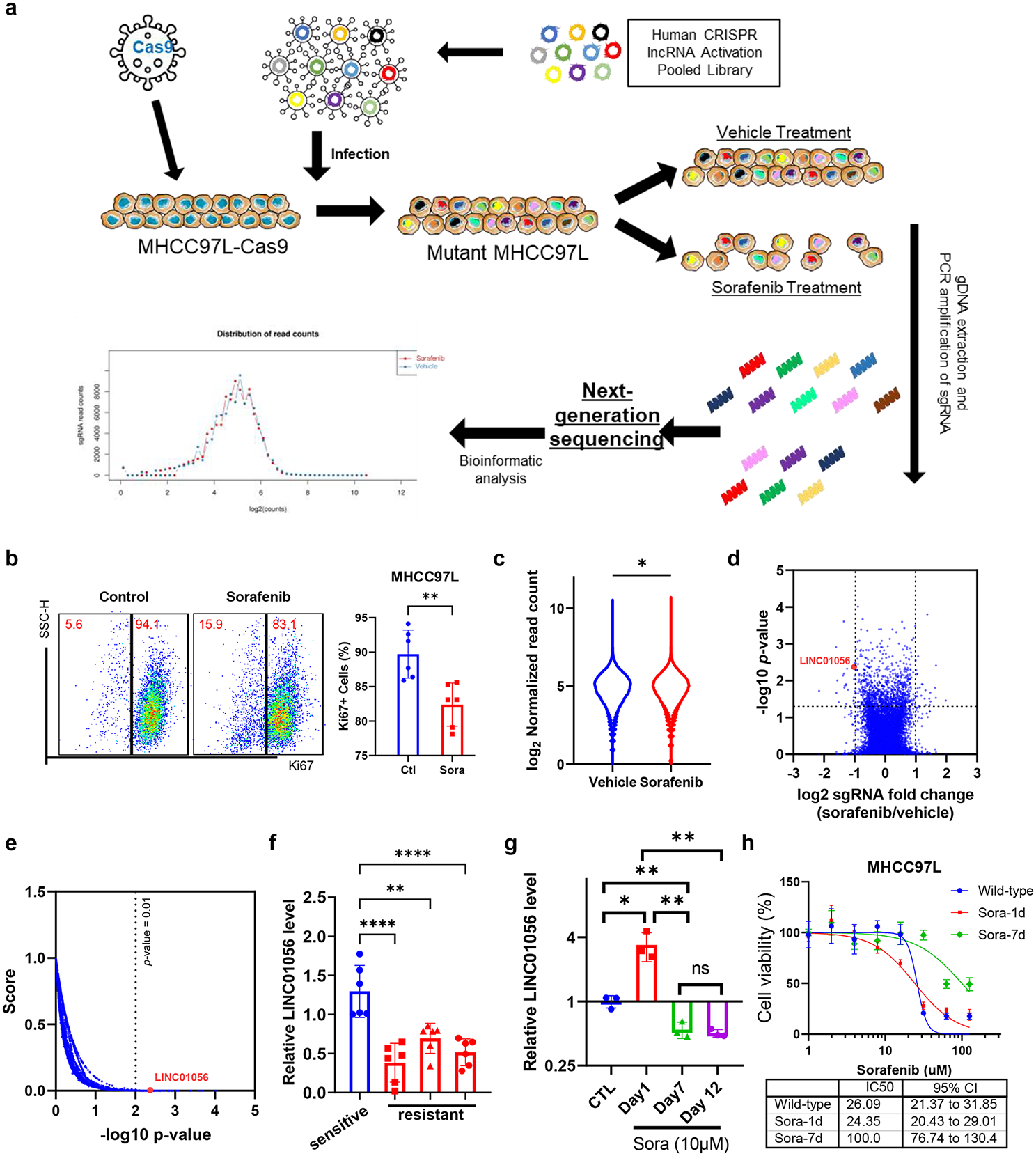
記住我
An 82-year-old woman was diagnosed in 2018 with invasive ductal carcinoma (IDC) of the right breast, grade 3, ER-/PR-/HER2 3 + , that had metastasized to the lymph nodes and bone. In 2019, contrast-enhanced brain computed tomography (CT) revealed a 3.9 cm cerebellar lesion that was surgically removed (Fig. 1A-C). We derived organoid cultures from this brain metastatic material (mOGs) by using a slightly modified organoid growth medium (OGM) containing neuregulin-1 to match the aberrant expression of ERBB family members in the starting material (Fig. 1D) [5, 6]. Using NGS-customized panels containing breast cancer-specific hotspot regions for 63 genes, we identified the PIK3CA H1047L mutation in both the patient-derived organoid (PDO) and the metastatic sample tissue, with variant allele frequencies (VAFs) of 45.4% and 56%, respectively (Fig. 1E and Suppl. Table 1). PIK3CA hotspot mutations drive oncogenic progression in several cancer settings [4]. These mutations occur in 30–40% of breast cancers and are associated with resistance to therapy and metastatic progression [7, 8]. We found that a very minor fraction of genes assessed in the mBC-PDOs exhibited different expression levels when compared with the brain metastasis sample of origin (Fig. 1F). This finding strongly validated the accuracy of the mBC-PDO model. Additionally, mutational analysis and Gene Set Enrichment Analysis (GSEA) of the paired metastatic lesion and the mBC-PDOs revealed that PI3K-related pathways were enriched (Fig. 1G).
Fig. 1
A Clinical features of case 1: primary breast cancer and relative brain metastases. B Contrast-enhanced brain computed tomography scan showing a solid and enhanced large lesion (3.9 cm) in the cerebellum with perilesional oedema. C Representative staining results from haematoxylin–eosin (HE) and immunohistochemistry for ER, PR, HER2 and Ki-67 in the metastatic lesion from patient 1. Scale bar = 200–500 µM. ER, oestrogen receptor; PR, progesterone receptor; HER2, human epidermal growth factor. D Representative bright-field microscopy image of organoids generated from metastatic lesions. Magnification: 20X. E Summary of the mutational profile of case 1 brain metastases and related organoids. VAF: variant allele frequency. F All expressed gene distributions by RNA-seq analysis of metastatic lesions (METs) and related organoids (PDOs). RNAseq data counts were normalized as transcripts per million (TPM). G Preranked gene set enrichment analysis (GSEA; https://www.gsea-msigdb.org) was performed on all expressed genes in the metastatic lesion (MET) and related organoid (PDO) samples. GSEA was run in preranked mode using classic as a metric and 1000 permutations selecting the curated gene sets of the Molecular Signatures Database (MsigDB) derived from the Hallmark and KEGG collections. As a ranking metric, the z scores of the genes were evaluated in each sample. Gene set enrichment was assessed through the normalized enrichment score (NES). The size of the circles reflects the percentage of genes in the core enrichment of the pathway. H Representative bright-field microscopy images of organoids generated from brain metastatic lesions carrying PIK3CA mutations treated with 10 µM alpelisib for 7 days. Magnification: 10X. The number of organoids and relative area for each condition are plotted in the bar graphs. I Flow cytometry of Helix NP blue-stained organoids generated from brain metastatic lesions carrying PIK3CA mutations after 7 days of treatment with 10 µM alpelisib. J Representative bright-field microscopy images of organoids generated from brain metastatic lesions with wild-type PIK3CA treated with 10 µM alpelisib for 7 days. Magnification: 10X. The number of organoids and relative area for each condition are plotted in the bar graphs. K Flow cytometry of Helix NP blue-stained organoids generated from brain metastatic lesions with wild-type PIK3CA after 7 days of treatment with 10 µM alpelisib
We first evaluated 105 genes representative of the PI3K pathway by RNAseq (MSigDB, https://www.gsea-msigdb.org) to assess the relationship between the mutation status of PIK3CA and the activation of the PI3K pathway. We found the levels of PI3K genes to be significantly higher in PIK3CA-mutant brain metastases than in PIK3CA-wild-type brain metastases; the latter sample was obtained from a 53-year-old woman diagnosed in 2019 with IDC of the right breast ER + /PR + /HER2- (Suppl. Fig. 1A and Suppl. Table 2). The difference in the levels of the PI3K-related genes was even more evident in the mPDO cultures (Suppl. Fig. 1B). We stained brain metastasis-derived PDOs with an anti-AKT-phosphoS473 antibody. The flow cytometry data analysis revealed an enrichment of AKT-phosphoS473-positive cells in the PIK3CA mutant PDOs compared with those in the PIK3CA wild-type PDOs. Altogether, these results suggest that the PI3K pathway was activated due to the experimental conditions (Suppl. Fig. 1C). Finally, a breast cancer TCGA cohort analysis further validated our results. In breast cancer samples bearing mutated PIK3CA, despite a nonsignificant modulation of the transcriptional levels of PI3K pathway genes, both AKT, AKT-phosphoS473, AKT-phosphoT308 and PRAS40-phosphoT246 protein levels were significantly increased compared to those in wild-type PIK3CA samples (Suppl. Fig. 1D-E).
To test the functional relevance of PI3K pathway activation in PIK3CA-mutant PDOs, we treated the organoids with alpelisib, a clinical trial grade PI3K-α inhibitor [9,10,11], at pharmacologically relevant doses. This compound affected the mBC-PDO number (images and graph) and size (area graph) and reduced the number of live cells in the PIK3CA-mutant mBC-PDOs, as assessed by flow cytometry (Fig. 1H-I). To further relate the efficacy of alpelisib treatment to the presence of the PIK3CA mutation, we similarly treated non-PIK3CA-mutated brain metastasis-derived PDO cultures (ctrl-mBC-PDO) with alpelisib. Treatment of the ctr-mBC-PDO cultures with 10 µM alpelisib had little to no effect on PDO number (images and graph) or size (area graph), and no changes in the number of live cells was observed by flow cytometry (Fig. 1J-K). Taken together, these findings further suggest the dependency of mBC-PDOs on aberrant PI3K-propagated signalling for their maintenance.
These encouraging findings prompted us to expand those observations to three additional cases of spine (case 2), lung (case 3) and skin (case 4) BC metastases, all with PIK3CA hotspot mutations. In detail, the case 2 sample was taken from a 61-year-old woman diagnosed in 2015 with pT2 pN3a IDC of the right breast, grade 3, ER + /PR + /HER2-, with a metastatic lesion to the spine (T4); the case 3 sample was taken from a 47-year-old woman diagnosed in 2014 with hormone-sensitive IDC breast cancer and multiple bilateral lung nodules, and the case 4 sample was taken from an 82-year-old woman with pT2 pN3 IDC of the left breast, grade 2, ER + /PR + /HER2-, with skin metastases on the left chest wall. All three patients had undergone previous Taxol and/or anastrozole treatment. The clinical and histopathological findings and representative magnetic resonance imaging (case 2) or computed tomography scans (cases 3 and 4) are shown in Fig. 2A. The receptor status, evaluated by immunohistochemistry, was ER + /PR-/HER2 2 + for the spine metastasis, ER + /PR + /HER2- for the lung metastasis and ER + /PR-/HER2- for the skin-derived metastasis (Fig. 2B), indicating changes in HR expression in the metastases versus that in the primary tumours. NGS tumour sequencing of the three metastatic lesions revealed the following PIK3CA hotspot mutations: E545K in case 2, H1047L in case 3 and H1047R in case 4 (Fig. 2C).
Fig. 2
A Clinical features of cases 2, 3 and 4 metastatic lesions and relative magnetic resonance imaging (case 2) or computed tomography scans (cases 3 and 4). B Representative staining results from haematoxylin–eosin (HE) and immunohistochemistry analyses of ER, PR, HER2 and Ki-67 expression in the metastatic lesions from patients 2, 3 and 4. Scale bar = 200–500 µM. ER, oestrogen receptor; PR, progesterone receptor; HER2, human epidermal growth factor. C Summary of the mutational profile of metastatic lesions of cases 2, 3 and 4. VAF refers to variant allele frequency. D Normalized enrichment score (NES) of the PI3K pathway among the four patient cases harbouring PIK3CA mutations (#1, #2, #3, #4) and the brain metastasis with an intact PIK3CA gene. All expressed genes were ranked after z score transformation, and enrichment analysis was conducted for each sample. The size of the circles reflects the percentage of genes in the core enrichment of pathway. E Representative bright-field microscopy images of organoids generated from metastatic lesions of patients 2 and 4 treated with 10 µM alpelisib for 7 days. Magnification: 10X. The number of organoids for each condition is plotted in the bar graph. F Flow cytometry of Helix NP blue-stained organoids generated from metastatic lesions of patients 2, 3 and 4 after 7 days of treatment with 10 µM alpelisib. G Number of live cells of organoids derived from metastatic lesions of patients 2 and 4 after 7 days of treatment with 10 µM alpelisib
We next analysed the bulk RNA-seq data of the metastatic lesions by comparing the normalized enrichment score (NES) of the PI3K pathway between the four cases harbouring PIK3CA mutations and the brain metastasis with no PIK3CA gene mutations to validate our previous findings. All the expressed genes were ranked after z score transformation, and enrichment analysis was conducted for each sample. As reported in Fig. 2D, the metastatic lesions harbouring PIK3CA mutations exhibited effective enrichment for the PIK3-driven pathway when compared to the PIK3CA wild-type lesions. We supplemented the basic OGM (neuregulin 7.5 nM, EGF 10 ng/mL, FGF2 50 ng/mL, B27, BSA 2%) with factors specific for the host tissue to generate mBC-PDO cultures from the PIK3CA-mutated metastases as follows: KGF (20 ng/mL) for skin metastases; FGF2 (50 ng/mL) for spine-derived metastases and an inverted ratio of FGF2 and EGF for lung-derived metastases (FGF2 20 ng/mL, EGF 40 ng/mL). The numbers of formed PDOs from the spine, skin and lung metastases were counted, revealing the effective formation of aggregates within 72 h after media addition. Additionally, the area of the formed structures was measured over time and showed that the structures were actively expanding (data not shown). Finally, flow cytometric analysis of the disaggregated PDOs confirmed that most of the cells composing the PDO cultures were viable (data not shown). We followed untreated PDOs up to passage 5, witnessing a small significant decrease in their growth over time (data not shown). Notably, no PDO structures grew when the metastatic lesions were cultured with the medium used to generate the brain metastasis PDOs.
The three newly obtained mBC-PDO cultures were then challenged with alpelisib, and a variable yet significant decrease in PDO number (graph) and size (area graph) was observed; additionally, the number of live cells was reduced in the spine- and skin-derived mBC-PDOs (Fig. 2E-G). Lung metastasis-derived PDOs showed the most limited response to alpelisib. This reduced effect could be partly due to the lower enrichment for PI3K pathway-related genes, despite the presence of PIK3CA mutation, as identified by NGS tumour sequencing (Fig. 2D). This finding echoes the current knowledge regarding the heterogeneity of metastatic lesions (Fig. 2F). Finally, we expanded this limited cohort of samples to include an additional sample derived from skin metastasis in an 86-year-old patient diagnosed with invasive ductal carcinoma (IDC) of the right breast, grade 3, ER + /PR-/HER2, harbouring the wild-type PIK3CA gene (Suppl. Fig. 2A). A viability assay performed on the PDO cultures obtained from this sample treated with the control (DMSO) or alpelisib revealed little to no PDO-derived cell death in response to the drug. This finding again strengthens the link between the response to alpelisib and the presence of a PIK3CA gene mutation (Suppl. Fig. 2B).
留言 (0)