
記住我
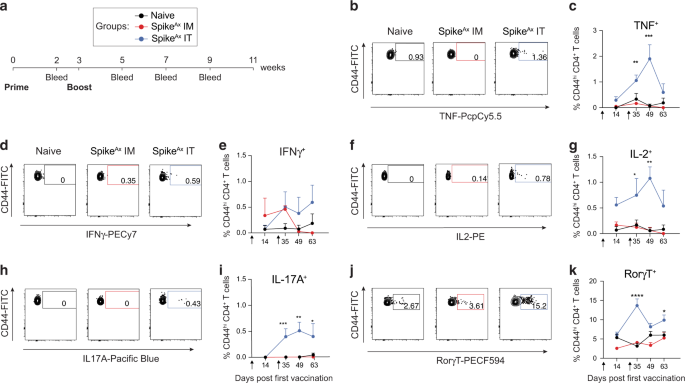

The respiratory tract is continuously exposed to airborne microbial products, which modulate the pulmonary immune system. Due to their strategic location in the alveoli of the lungs, AMs are in direct contact with inhaled particles and microbes, and thus represent potential candidates to develop trained immunity. To investigate whether ambient concentrations of ubiquitous airborne compounds can elicit AM memory, we administered 1 ng LPS (∼0.5 EU/mouse; an amount in relation potentially inhaled by humans8) or endotoxin-free saline intranasally (i.n.) to wild type C57BL/6 J mice (Fig. 1a). This treatment induced an acute inflammatory response, characterized by a transient influx of neutrophils after 24 h (Fig. 1b). Six days later, neutrophils were no longer detectable in the BAL fluid (BALF; Fig. 1c), and post-lavage lung immune cell numbers were comparable to the control group, except for dendritic cell and B cell numbers, which remained moderately elevated in LPS-exposed lungs (Fig. S1a–c). These data support the transient nature of LPS-triggered effects, and exclude overt signs of persisting lung inflammation. To assess whether LPS exposure modulates AM immunity, we isolated AMs by BAL six days after i.n. LPS treatment and challenged the cells ex vivo with heat-inactivated S. pneumoniae (HISP, Fig. 1a). LPS-exposed AMs produced higher amounts of multiple cytokines and chemokines, including C-X-C Motif Chemokine Ligand (CXCL)-1, interleukin (IL)-1β, IL-10, IL-12p40 and IL-6 compared to control (saline-exposed) AMs (Fig. 1d, Fig. S2a), indicating an innate memory effect. Taking absolute concentrations and log2 fold changes into account (Fig. 1d), we selected IL-6, a cytokine critically involved in host immunity16, as the most stable readout and decided to use it as a surrogate for LPS-induced AM memory in subsequent experiments. In support of this choice, we continued to observe elevated IL-6 responses by AMs two and six weeks after intranasal LPS treatment (Fig. 1e), indicating long-lasting cellular reprogramming following ambient LPS exposure.
Fig. 1: LPS exposure induces trained immunity in alveolar macrophages.
a Experimental setup for i.n. LPS (1 ng/mouse) or saline exposure, followed by FACS analysis (BALF, lung) or AM cytokine analysis upon ex vivo bacterial challenge. b, c Absolute numbers of BALF AMs and neutrophils, measured by flow cytometry 24 h (b) and six days (c) after in vivo treatment. d LEGENDplex analysis of LPS-exposed and control AMs upon ex vivo HISP-challenge (16 h). Absolute cytokine levels (top heat map) and log2 fold change (log2fc) of LPS-exposed AMs versus means of control AMs (bottom heat map) are shown. The right panel compares absolute levels and log2fc of trained AMs. e IL-6 levels of LPS-exposed and control AMs upon ex vivo bacterial challenge two or six weeks after treatment. f Phagocytosis index (AU: arbitrary units) of trained and control AMs, isolated on day six after in vivo training, followed by ex vivo stimulation with FITC-labeled HISP and FACS analysis. g Experimental setup for PKH26 labeling eight days prior to in vivo training. h, i Representative histograms of PKH26 MFI (gated on CD11c+ Siglec F+ AMs) and percentage of PKH26+ AMs 24 h (h) and six days (i) after training. Graphs show means + SD of 7 (b) or 11–12 (c, h, i) biological replicates, means of 7–8 biological replicates (d) or means + SD of 3–5 technical replicates (e, f). Data (b, c, e, f) are representative of two independent experiments. Statistical analysis: student’s t-test. ns, not significant. *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001, ****p ≤ 0.0001.
Given that phagocytosis and efferocytosis are key effector functions of AMs, we next decided to investigate whether these processes are modulated six days after in vivo training. To analyze AM-mediated phagocytosis, AMs were isolated by BAL and incubated with FITC-labeled HISP, followed by FACS analysis. These experiments revealed that the phagocytic capacity of trained AMs was enhanced in comparison to saline-treated controls (Fig. 1f). AM-mediated efferocytosis was assessed after intratracheal transfer of CFSE-labeled apoptotic thymocytes on day six after training (Fig. S2b) and did not reveal any differences between the groups (Fig. S2c, d). However, we noticed that LPS-exposed AMs showed elevated surface expression of MerTK (Fig. S2e) and Axl (Fig. S2f), two TAM family receptors known to promote apoptotic cell removal17.
AMs are predominantly of embryonic origin and self-maintain locally with minimal contribution of circulating monocytes under steady-state conditions18. However, upon infection or severe lung injury, the alveolar niche can be replenished by monocytes, which get recruited to the lungs and acquire an AM profile under the influence of the local microenvironment13. In order to determine whether the transient inflammatory response to LPS inhalation induced replenishment of tissue-resident AMs by monocytes, we labeled resident AMs by i.n. administration of the fluorescent dye PKH2619 eight days prior to LPS treatment (Fig. 1g). Frequencies of PKH26+ and PKH26− AMs were determined 24 h and six days after training. The dye selectively labeled lung-resident CD11c+ Siglec F+ AMs while CD11b+ Ly6C+ monocytes remained PKH26-negative (Fig. S2g). Of note, the percentage of PKH26+ CD11c+ Siglec F+ AMs in BALF (Fig. 1h, i) and post-lavage lung tissue (Fig. S2h) was comparable between LPS-treated and control mice at both timepoints investigated. This indicates that the resident AM population was not replenished by inflammatory monocytes upon in vivo training. In conclusion, we showed that i.n. LPS exposure trains the local AM pool for increased cytokine production and phagocytosis following secondary bacterial challenge.
AM training depends on type 1 interferon signalingLPS-mediated signaling is accompanied by the production of type 1 interferons (IFNs; e.g. IFN-β)20 and type 2 IFNs (IFN-γ)21, which have the capacity to modulate immune responses. Yao et al. reported that IFN-γ, produced by T-cells during respiratory adenoviral infection, primes AMs for enhanced immune activity upon secondary pneumococcal challenge22. To test whether IFN-γ- or T cell-mediated responses contribute to LPS-induced AM memory, we applied our training model (in vivo training, followed by ex vivo AM challenge) to IFN-γ-receptor-deficient (Ifngr1−/−) and Rag2-deficient (Rag2−/−) mice, respectively. These experiments showed that the enhanced IL-6 response of LPS-exposed AMs occurred independently of IFN-γ-receptor signaling (Fig. S3a) and adaptive immunity (Fig. S3b).
LPS is a potent inducer of type 1 IFNs, which can in turn potentiate LPS-mediated immune responses23. To investigate whether type 1 IFN signaling plays a role in LPS-induced AM memory, we trained type 1 IFN-receptor-deficient (Ifnar1−/−) and control mice, and analyzed AM IL-6 production upon ex vivo HISP challenge six days later. Interestingly, AMs retrieved from LPS-exposed Ifnar1−/− mice were unable to mount a trained response (Fig. 2a), suggesting that type 1 IFN signaling plays an important role in the establishment of AM memory. Of note, this effect was not restricted to IL-6, as IL-12p40 and IL-12p70 responses were similarly diminished upon Ifnar1 deficiency (Fig. S3c, d).
Fig. 2: LPS-induced AM training is driven by type 1 interferon signaling.
a, b IL-6 levels of HISP-challenged (16 h) LPS-trained and control AMs, isolated six days after in vivo training (performed as described in Fig. 1) of Ifnar1−/− and WT control mice (a) or Ifnar1ΔCD169 and Ifnar1fl/fl control mice (b). c Experimental setup for i.n. treatment with IFN-β (2000 U/mouse) or saline (control). AMs were isolated on day six, followed by ex vivo challenge with HISP (16 h). d IL-6 levels of IFN-β-trained and control AMs after ex vivo restimulation. e Experimental setup for in vitro LPS (10 ng/mL)- or IFN-β (800 U/ mL)- training of mexAMs, followed by in vitro HISP challenge (16 h) six days later. f IL-6 levels of control, LPS-trained and IFN-β-trained WT mexAMs after HISP challenge. g IL-6 levels of LPS-trained and control Ifnar1ΔCD169 and Ifnar1fl/fl mexAMs after HISP challenge. Graphs show means + SEM of 4-5 technical replicates. In (a–d), four biological replicates per group were pooled and seeded as technical replicates for stimulation. Data are representative of two independent experiments. Statistical analysis: two-way ANOVA (factor 1: training; factor 2: genotype) (a, b, g), student’s t-test (d) or one-way ANOVA (f). ns, not significant. *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001, ****p ≤ 0.0001.
Based on these findings, we aimed to dissect whether type 1 IFNs promote AM training in a direct or indirect manner. Using Ifnar1ΔCD169 and Ifnar1fl/fl control mice, we found that macrophage-specific deficiency of type 1 IFN receptor expression abolished AM memory (Fig. 2b), indicating that AMs need to directly sense type 1 IFNs during LPS-mediated training. This prompted us to investigate whether local administration of type 1 IFNs has the potential to induce trained immunity in AMs. For this purpose, we treated wild type mice i.n. with IFN-β or saline and examined the responsiveness of AMs six days later (Fig. 2c). Similar to LPS-training, IFN-β exposure increased the IL-6 production by AMs upon secondary, bacterial challenge, indicating an innate memory effect (Fig. 2d).
Given that type 1 IFNs act downstream of TLR4 signaling and can affect cellular immunity via autocrine signaling24, we went on to explore whether AM memory can be induced in a cell-autonomous manner and generated murine ex vivo cultured AMs (mexAMs)25 from primary wild type AMs. Following in vitro expansion, mexAMs were stimulated for 24 h with LPS, IFN-β or medium, allowed to rest for five days, and subsequently challenged with HISP on day six after training (Fig. 2e). Similar to in vivo AM training, LPS- or IFN-β-exposed mexAMs produced increased amounts of IL-6 upon bacterial challenge (Fig. 2f), indicating that these stimuli have the potential to directly induce AM memory. Applying the same regimen to Ifnar1ΔCD169 and Ifnar1fl/fl control mexAMs, we could further demonstrate that autocrine type 1 IFN signaling can mediate LPS-induced in vitro training (Fig. 2g).
Collectively, our data suggest that type 1 IFNs play an important role in the establishment of LPS-mediated AM memory.
Trained AMs exhibit an altered transcriptional profile upon secondary bacterial challengeTrained immunity is defined as the altered reactivity to a secondary trigger induced by prior exposure to a training stimulus26. In contrast to primed immune responses, this phenomenon is characterized by the return to a baseline state after initial activation26. Mechanistically, trained immunity has been associated with epigenetic remodeling and metabolic reprogramming, two processes that serve as the molecular basis for altered gene expression upon secondary challenge3. To identify transcriptional changes of LPS-exposed and control AMs at baseline and upon subsequent bacterial stimulation, AMs were isolated six days after training and incubated for 3 h with medium or HISP (Fig. 3a). Principal component analysis (PCA) of RNA-seq results revealed a high similarity between trained and control AMs (Fig. 3b, c), and only 10 differentially expressed genes (DEGs) at baseline (Table S1). In contrast, transcriptional profiles clustered according to the preceding training stimulus upon bacterial challenge (Fig. 3b, c), which correlated with 165 upregulated and 27 downregulated genes identified in LPS-trained compared to control AMs (Table S2). DEGs detected upon HISP challenge mapped to different Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways (Fig. 3d), with “cytokine-cytokine receptor interaction“ and “chemokine signaling“ being most differentially regulated. Among these pathways, signaling-related genes (Prkcd, Nfkb1, Raf1) as well as genes encoding chemokines of CC (Ccl22, Ccl3) and CXC (Cxcl2, Cxcl3) subfamilies were differentially expressed upon HISP stimulation (Fig. 3e). In accordance, CXCL1 protein levels were markedly increased upon ex vivo challenge of trained AMs (Fig. S2a).
Fig. 3: LPS-trained AMs display an altered gene expression profile upon ex vivo challenge.
a Experimental setup for ATAC-seq and RNA-seq analysis of AMs isolated on day six after in vivo training. b Principal component analysis of normalized gene expression data obtained from in vivo trained AMs upon 3 h ex vivo HISP challenge (squares) or medium (circles). c Heatmap depicting DEGs in trained and control AMs upon medium or HISP challenge. Samples are clustered by unsupervised clustering. Data are rlog transformed, followed by z-score scaling. Cutoff: adjusted p-value ≤ 0.1; red horizontal bars adjacent to heatmaps indicate statistical significance. d Top 20 KEGG pathways in HISP-challenged trained and control AMs. Circle size indicates the number of DEGs associated with the respective pathway. e Heatmaps depicting DEGs identified in the top two differentially regulated pathways in trained and control AMs upon medium or HISP challenge. Data are rlog transformed, followed by z-score scaling. f Volcano plot displaying differentially accessible regions (DARs; padj ≤ 0.05) of trained versus control AMs identified by ATAC-seq analysis six days after in vivo training. Labels indicate top 10 DARs per group.
To assess whether the altered transcriptional responsiveness of LPS-exposed AMs was associated with persistent epigenetic changes reflected by altered chromatin accessibility, BAL AMs were processed for Assay for Transposase-Accessible Chromatin (ATAC)-seq analysis on day six after in vivo training (Fig. 3a). In total, we identified 24 differentially accessible regions (DARs; Fig. S4a and Table S3; FDR ≤ 0.05), nine of which were more accessible in the trained group (Fig. 3f). Among these, three DARs were annotated to the genes Fos (2 DARs) or Fosb (1 DAR), which are associated with transcriptional regulation of multiple biological processes, including cell migration, differentiation and inflammation27.
Next we considered the possibility that accelerated gene expression upon secondary challenge may result from altered baseline deposition of permissive histone marks, such as H3K4 methylation or H3K27 acetylation28. To test whether AM training depends on methylation- or acetylation events established during LPS exposure, we trained WT mexAMs in presence of the methyltransferase inhibitor 5’-deoxy-5′-methylthioadenosine29 (MTA), the acetyltransferase inhibitor anacardic acid30 or DMSO. After 24 h, cells were washed and allowed to rest in medium until bacterial challenge (Fig. S4b). Neither of the inhibitors influenced the trained IL-6 response (Fig. S4c), suggesting that the targeted epigenetic enzymes do not contribute to LPS-induced AM memory.
Collectively, trained and control AMs displayed similar gene expression levels and few changes in chromatin accessibility at baseline, but mounted an augmented transcriptional response upon secondary, bacterial challenge.
Secondary metabolic AM responses are modulated by prior LPS exposureRecent research highlighted the critical impact of cellular metabolism on the functional state of immune cells, including the induction, maintenance and regulation of trained immunity31. Importantly, metabolic pathways do not only provide energy and macromolecular building blocks, but can directly influence the epigenetic machinery by generating intermediate metabolites that serve as substrates or cofactors32. In monocytes and macrophages, glycolysis, glutaminolysis and cholesterol synthesis have been described to play crucial roles in the induction of trained immunity, with increased glycolysis defined as hallmark of trained macrophages31,32. AMs, however, are unique tissue-resident immune cells that are metabolically adapted to the remarkably low glucose concentration of the alveolar space, and primarily utilize OXPHOS to meet their energy demands14. We therefore speculated that the metabolic characteristics of trained AMs may differ from those classically associated with trained monocytes and macrophages. To investigate the baseline metabolism of AMs six days after in vivo training, we performed Seahorse analyses (Fig. 4a, b). Compared to control AMs, LPS-exposed AMs displayed a reduced basal metabolic activity, reflected by a decreased oxygen consumption rate (OCR) and extracellular acidification rate (ECAR) (Fig. 4c). Sequential inhibition of selected electron transport chain (ETC) components further revealed a reduced ATP production rate of trained AMs (Fig. 4c), while maximum respiratory capacity and spare respiratory capacity (SRC) were unaltered (Fig. S5a). To assess how prior LPS exposure affects AM metabolism upon secondary, bacterial challenge, we performed Seahorse analyses of trained and control AMs 16 h after incubation with HISP (Fig. 4d, e). Interestingly, basal OCR and ECAR, ATP production (Fig. 4f), as well as maximum and spare respiratory capacity (Fig. S5b) were significantly decreased in trained cells. While saline-exposed AMs exhibited an increased OCR upon bacterial stimulation (16 h; compared to incubation with medium only), trained AMs displayed similar OCR levels in presence or absence of HISP (Fig. 4g). Although both trained and control cells increased their ECAR upon bacterial challenge (Fig. 4g), the absolute ECAR of trained AMs was lower than that of control AMs at the investigated time point.
Fig. 4: LPS exposure persistently alters the metabolic state and responsiveness of AMs.
a Experimental setup for Seahorse analyses of AMs on day six after in vivo training with LPS/saline. b OCR and ECAR of trained and control AMs, measured at baseline and after sequential treatment with oligomycin (Oligo), FCCP and rotenone/antimycin A (R/A). c Quantification of baseline OCR (basal respiration), ATP production rate and baseline ECAR. d Experimental setup for Seahorse analyses of AMs upon ex vivo HISP or medium challenge (16 h) on day six after training. e OCR and ECAR of trained and control AMs, 16 h after HISP stimulation, measured at baseline and after sequential treatment with indicated drugs. f Quantification of basal respiration, ATP production rate and baseline ECAR 16 h after HISP challenge. g Quantification of baseline OCR and ECAR 16 h after stimulation with HISP in medium versus medium only. Graphs show means + SEM of 10-11 technical replicates from pooled biological replicates (n = 5-8). Data are representative of two independent experiments. Statistical analysis: student’s t-test. ns, not significant. ***p ≤ 0.001, ****p ≤ 0.0001; OCR, oxygen consumption rate; ECAR, extracellular acidification rate; FCCP, carbonyl cyanide-4-(trifluoromethoxy)phenylhydrazone.
In summary, these experiments demonstrate that ambient LPS exposure rewires AM metabolism, which further impacts the metabolic response to subsequent bacterial challenge.
LPS training induces changes in AM metabolite and lipid compositionBased on these findings, we went on to investigate the effects of LPS training on the metabolite and lipid composition of AMs. CD11c+ Siglec F+ AMs were FACS-sorted six days after LPS administration and subjected to LC-MS/MS-based analyses (Fig. 5a; Fig. S6a). Targeted metabolomic analysis revealed that trained AMs contained increased amounts of S-adenosyl-methionine (SAM; Fig. 5b), an essential metabolite synthesized from methionine and ATP33. In accordance, intracellular amino acid profiles (Fig. S6b) showed a trend for increased methionine concentrations in trained AMs compared to control cells (Fig. S6c). SAM acts as a universal methyl group donor for RNA, DNA, lipids and proteins and was reported to drive a proinflammatory macrophage phenotype in the context of LPS-induced inflammation34. In addition, LPS exposure modulated metabolites of the tricarboxylic acid (TCA) cycle, with fumarate and malate being significantly reduced compared to control AMs (Fig. 5b). Interestingly, lipidomic analysis identified profound differences between LPS- and saline-exposed AMs on day six after training (Fig. 5c). Trained AMs contained substantially higher amounts of selected ceramides (Cer), phosphatidylethanolamines (PE), sphingomyelins (SM) and phosphatidylcholines (PC), essential membrane lipids that can directly or indirectly impact membrane receptor signaling35. In contrast, triacylglycerol (TAG) levels were strongly reduced in trained compared to control AMs (Fig. 5d). TAGs serve as cellular energy stores that fuel cell-intrinsic ATP production in the mitochondria by providing free fatty acids for β-oxidation36. In summary, these findings imply that in vivo LPS exposure profoundly modulates the metabolite and lipid composition of AMs.
Fig. 5: LPS-induced metabolic activation contributes to the establishment of AM memory.
a Experimental setup for metabolomic/lipidomic analyses of AMs on day six after in vivo training with LPS/saline. b AM intracellular metabolites related to the tricarboxylic acid (TCA) cycle. SAM: S-adenosyl-methionine. c Principal component analysis of centered log-ratio transformed AM lipidomics data. d Heatmap displaying differentially abundant lipids (FDR 0.2, p-value ≤ 0.05) of trained and control AMs. Metabolomic/lipidomic analyses were performed with 5-6 biological replicates per group. Cer ceramides; DAG diacylglycerols; PE phosphatidylethanolamines; SM sphingomyelins; PC phosphatidylcholines; CE cholesterol esters; TAG triacylglycerols. e Experimental setup for mexAM training with LPS or medium in presence of indicated metabolic inhibitors or DMSO, followed by in vitro HISP challenge (16 h) six days later. f IL-6 levels of mexAMs stimulated as described in e. Data are representative of two independent experiments. Graphs show means + SD of 5-6 biological replicates (b) or means + SEM of 4-5 technical replicates (f). Statistical analysis: student’s t-test (b) and two-way ANOVA (f) (factor 1: training; factor 2: inhibitor). ns, not significant. *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001, ****p ≤ 0.0001.
LPS-induced rewiring of AM metabolism is critical for memory inductionTissue-resident AMs predominantly rely on OXPHOS to meet their metabolic demands37. This process is tightly linked to the TCA cycle, which serves as the main electron donor for the mitochondrial ETC. Substrates fueling the TCA cycle can be generated by multiple processes, including fatty acid oxidation (FAO), glutaminolysis and oxidation of amino acids or pyruvate38. Based on the altered metabolite and lipid composition of LPS-exposed AMs, we speculated that the initial metabolic activation evoked by the training stimulus might be critical for the altered reactivity observed upon secondary challenge. To test this hypothesis, we applied selective metabolic inhibitors during in vitro mexAM training and determined the consequences on the trained IL-6 response exerted upon bacterial challenge. 2-deoxyglucose (2-DG), bis-2-(5-phenylacetamido-1,3,4-thiadiazol-2-yl)ethyl sulfide (BPTES) and etomoxir were used to inhibit glycolysis, glutaminolysis or FAO, respectively (Fig. 5e). While glycolysis appeared to be dispensable for the establishment of mexAM memory, inhibition of FAO and glutaminolysis abrogated the trained IL-6 response of LPS-exposed cells (Fig. 5f), suggesting that LPS-mediated metabolic rewiring is critical for the establishment of AM memory.
LPS training modulates pneumonia outcomeAM activation represents an ambiguous balancing act, which serves to promote pathogen clearance while maintaining tissue integrity. Consequently, malfunction, hypo-or hyperactivation of AMs can have detrimental consequences for the host12,39. To investigate whether LPS-trained AMs can modulate the outcome of a subsequent pneumococcal infection, we isolated AMs five days after in vivo training (Fig. S7a–c) and transferred them intratracheally (i.t.) into naïve recipients (Fig. S7d), followed by i.n. S. pneumoniae infection (“in vivo challenge”) 24 h later (Fig. 6a). Recipients of trained AMs displayed increased bacterial loads (Fig. 6b) and lung inflammation (Fig. 6c, d), indicating that adoptive transfer of LPS-experienced AMs impairs host defense against bacterial pneumonia. Considering that LPS-mediated effects on other (i.e. non-AM) cell populations are omitted in a transfer setup, we next addressed the impact of LPS training in a physiologically more relevant setting and infected mice with S. pneumoniae six days after LPS exposure (Fig. 6e). These experiments revealed that LPS-pretreated animals displayed enhanced bacterial clearance (Fig. 6f) and reduced lung tissue inflammation (Fig. 6g, h) 48 h after infection. While LPS-exposed animals demonstrated increased recruitment of inflammatory cells early upon infection (6 h; Fig. S7e–f), monocyte and neutrophil numbers were decreased after 48 h (Fig. S7h), indicating accelerated initiation and resolution of inflammation.
Fig. 6: LPS exposure modulates pneumonia outcome in a context-dependent manner.
a Experimental setup for adoptive transfer of LPS-trained or control donor AMs, followed by i.n. S. pneumoniae infection (in vivo challenge). Donor AMs were isolated by BAL five days after in vivo training and transferred intratracheally (i.t.) to naïve WT mice. Recipients were i.n. infected with S. pneumoniae 24 h after cell transfer. b Lung bacterial loads of recipients, determined 48 h after infection. c, d Representative histology images (c) and pneumonia score (d) of H&E-stained lung tissue 48 h after infection; scale bars: 100 µm. Graphs show means + SEM of two pooled experiments with 6–8 biological replicates each (total n = 12–16). e Experimental setup for in vivo training, followed by infection with S. pneumoniae (in vivo challenge) on day six. f Lung bacterial loads, 48 h after infection. g, h Representative histology images (g) and pneumonia score (h) of H&E-stained lung tissue, 48 h after infection; scale bars: 200 µm. Graphs show means + SD of 8–10 biological replicates. Data are representative of two independent experiments. Statistical analysis: Mann-Whitney-U test. *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001.
Overall, these results highlight the necessity to investigate the physiological consequences of environmental exposures as they may be influenced by multiple cellular players and tissue-specific parameters.
留言 (0)