
記住我
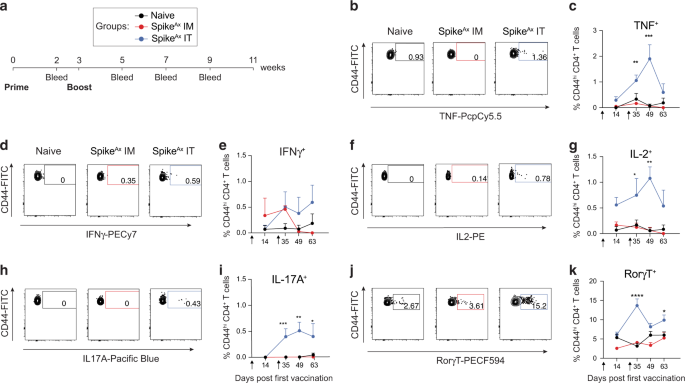

To investigate the role of NmU in asthma, we first addressed the question of whether NmU-25 is detectable in human samples. Blood and induced sputum were collected from healthy donors and patients with asthma. The concentration of hNmU-25 in plasma and sputum supernatants was determined with enyme-linked immunosorbent assay (ELISA) (Fig. 1a). hNmU-25 was detectable in both plasma and sputum supernatants, with slightly higher concentrations in the sputum. However, no significant difference in concentrations was detected between samples from healthy controls and patients with asthma.
Fig. 1: Expression of NmU and NmUR1 in human samples.
a hNmU-25 levels in human plasma and sputum supernatants from healthy subjects or patients with asthma were detected by ELISA. b Comparison of NmUR1 expression in different human immune cells from fresh blood determined with flow cytometry. c NmUR1 expression in CRTH2+ (type 2 lymphocytes: Th2, Tc2 and ILC2) and CRTH2- lymphocytes (other Th, Tc, and ILC). The expression levels in type 2 cells were compared with gMFI. d, e NmUR1 expression in ILC subsets (d) or in ILC2 subsets (e). f–h NmUR1 expression in Th2 (f), Tc2 (g) and ILC2 (h) cells in fresh blood among healthy controls (healthy), non-eosinophilic (non-eos) and eosinophilic (eos) asthma groups was compared. *p < 0.05, **p < 0.01, ***p < 0.001, ***p < 0.0001.
NmUR1 is expressed by human type 2 lymphocytes and upregulated in lung T cellsWe next investigated the expression of NmU receptors in human immune cells from fresh blood (Fig. 1b). NmUR1 was detected in most immune cell types except neutrophils by flow cytometry. More NmUR1 positive cells were found in type 2 immune cells including Th2, Tc2, ILC2 cells and eosinophils. In contrast, NmUR2 was undetectable in these cells (Supplementary Fig. 1a).
In lymphocytes, NmUR1 expression was higher in the CRTH2 positive type 2 compartment compared to CRTH2 negative counterparts, although there was no significant difference among Th2, Tc2 and ILC2 cell types (Fig. 1c; Supplementary Fig. 2). In ILCs, NmUR1 expression was similar in ILC1s and ILC2s, while slightly lower in ILC3s (Fig. 1d). We also compared NmUR1 expression in different subsets of ILC2s22,23, but no significant difference was detected (Fig. 1e).
To investigate the expression of NmUR1 in asthma, we compared the levels of NmUR1 in type 2 lymphocytes between healthy individuals and patients with asthma (Supplementary Table 1). The expression levels of the receptor, in terms of the percentage of positive cells (Fig. 1f–h upper panels) and geometric mean fluorescence intensity (gMFI) (Fig. 1f–h lower panels), were similar between healthy controls, non-eosinophilic asthma and eosinophilic asthma groups.
We further compared the expression levels of NmUR1 in type 2 cells between the peripheral blood and the airways by using paired blood and resected lung tissues (Fig. 2). Flow cytometric analysis revealed that the levels of NmUR1 in lung-derived Th2 and Tc2 cells were higher than those in blood-derived samples (Fig. 2a, b). Expression levels in lung-derived ILC2s were comparable to those in blood. Similar to blood, the expression of NmUR1 in CRTH2+ type 2 compartments was significantly higher than that in CRTH2- counterparts (Fig. 2c), although the level of NmUR1 in lung-derived CRTH2- Th cells was also increased compared with blood (Supplementary Fig. 3a). To further investigate the mechanism of NmUR1 upregulation in the lung, we compared T cells derived from blood and lung tissues by staining the cells with tissue residency markers CD69 and CD103. The ratio of CD69+ T cells and CD69+CD103+ T cells was significantly increased in CD4+ and CD8+ T cells from lung (Supplementary Fig. 3b). The increase of NmUR1 positive cells in lung tissues appeared to be mainly attributable to CD69+ T cells, as the most significant enrichment of NmUR1+ Th2 and Tc2 cells detected in lung mononuclear cells (LMCs) was in CD69+ cells (Fig. 2d, e) but not CD103+ or CD69+CD103+ cells, although the proportion of all three groups (CD69+, CD103+ or CD69+CD103+) of cells in total NmUR1+ type 2 T, Th2 or Tc2 cells was increased in lung samples (Fig. 2 f). To determine whether the upregulation of NmUR1 in the lung tissues was due to cell activation, we also stained the cells from the lung tissues with cell activation markers CD38, CD45RA and HLA-DR. The levels of CD69 in NmUR1+ T cells in the lung were not associated with those of activation markers (Supplementary Fig. 4a, b), although the levels of CD38 and HLA-DR are slightly higher in CD69+NmUR1+ cells compared with NmUR1- counterparts (Supplementary Fig. 4c). This suggests the upregulation of NmUR1 in the lung-resident memory type 2 T cells was not purely due to cell activation. However, we also observed an increase of CD38+ and HLA-DR+ cells and decrease of CD45RA+ cells in NmUR1+ T cells from the lung compared to blood (Fig. 2g). Although the latter changes were not associated with CD69 expression (Fig. 2h), these data taken together indicate that cell activation may contribute in part to the upregulation of NmUR1 in the lung.
Fig. 2: Expression of NmUR1 is upregulated in type 2 cells from lung.
a Histograms and b individual value plots to compare NmUR1 expression in Th2, Tc2 and ILC2 cells from PBMCs and LMCs detected with flow cytometry. c Comparison of NmUR1 expression in CRTH2+ (Th2, Tc2 and ILC2) and CRTH2- (other Th, Tc, and ILC) lymphocytes from lung tissues. d Increase of NmUR1+ cells in type 2 T cells from lung samples was mainly contributed by CD69+ cells. e Histograms to show the ratios of CD69 positive cells in different NmUR1+ type 2 cell groups. f Comparison of frequencies of CD69+, CD103+ or CD69+CD103+ Th2 or Tc2 cells in total NmUR1+ type 2 T cells between blood and lung. g CD38+ and HLA-DR+ cells were increased and CD45RA+ cells were decreased in the NmUR1+ T cells from LMCs. h The upregulation of CD38 and HLA-DR in the lung was mainly in CD69- but not CD69+ cells. a, d and e are representative of 7 (a) or 6 (d and e) independent experiments. *p < 0.05, **p < 0.01 and ***p < 0.001.
hNmU-25 induces human type 2 lymphocyte migrationTo explore the role of NmU in human type 2 immunity, Th2, Tc2, and ILC2 cells were isolated from human blood and cultured for further in vitro investigation (Supplementary Fig. 2b). NmUR1 expression was detected in these cultured type 2 cells at both transcriptional (mRNA) and protein levels (Fig. 3a, b). In contrast, the expression of NmUR2 in the cultured type 2 cells was negligible (Fig. 3a and Supplementary Fig. 1b, c). No significant difference in expression levels of NmUR1 was observed among Th2, Tc2, and ILC2s (Fig. 3c), although higher mRNA level of NmUR1 was detected in cultured ILC2s (Fig. 3a).
Fig. 3: NmUR1 is expressed in cultured human type 2 lymphocytes.
a mRNA levels of NmUR1 and NmUR2 in type 2 cells were compared with qRT-PCR. EEF1A1 was used as a control gene. b Expression of NmUR1 in type 2 cells was detected with Western blot. β-actin was used as a control protein. c NmUR1 expression in type 2 lymphocytes was compared with flow cytometry. ***p < 0.001, ****P < 0.0001, (n = 4–8 for a; n = 3 for b).
To understand whether the NmUR1 in these cultured type 2 lymphocytes was biologically functional, the Th2, Tc2, and ILC2 cells were treated with dose titrations of hNmU-25 in chemotaxis assays (Fig. 4a). hNmU-25 induced cell migration in a dose-dependent manner, exhibiting typical bell-shaped curves of chemotaxis peaking at approximately 1 µM in Th2 and ILC2s, and 100 nM-1 µM in Tc2 cells.
Fig. 4: NmU induces activation of type 2 lymphocytes.
a Th2, Tc2 and ILC2 cell migration to hNmU-25 measured with chemotaxis assays. b, d, f mRNA levels of IL5 and IL13 in cultured Th2 (b), Tc2 (d) and ILC2 (f) cells after treatment with various concentrations of hNmU-25 for 4 h detected with qRT-PCR. GAPDH was used as a control gene. c, e, g Protein levels of IL-5 and IL-13 in the supernatants of Th2 (c), Tc2 (e) and ILC2 (g) cultures after treatment with various concentrations of hNmU-25 for 4 h measured with ELISA. h, i Levels of CD38 (h) and HLA-DR (i) in CRTH2- or CRTH2+ T cells in fresh PBMCs treated with 100 nM hNmU-25 determined with flow cytometry. *p < (n = 5–7 for a; n = 3–4 for b, d and f; n = 3–5 for c, e and g; n = 10 for h, i).
hNmU-25 induces type 2 cytokine production in human type 2 lymphocytesTo further study the role of NmU in the activation of human type 2 lymphocytes, cultured cells were treated with serial concentrations of hNmU-25 for analysis of cytokine production. The transcriptional levels of IL5 and IL13 in the cells were measured by quantitative RT-PCR (qRT-PCR) (Fig. 4b, d, f), and the protein levels of IL-5 and IL-13 released in the cell supernatants were measured by ELISA (Fig. 4c, e, g). The treatment increased IL-5 and IL-13 levels at both transcriptional and protein levels in all three types of cells in a dose-dependent manner. The EC50 of hNmU-25 for IL5 and IL13 mRNA up-regulation in Th2 cells was 3.5 and 4.5 nM, in Tc2 cells was 7.25 and 3.4 nM, and in ILC2s was 9.27 and 0.8 nM, respectively (Fig. 4b, d, f). The EC50 of hNmU-25 for IL-5 and IL-13 production in Th2 cells was 4.57 and 44.5 nM, in Tc2 cells was 2.68 and 7.33 nM, and in ILC2s was 84 and 3.2 nM, respectively (Fig. 4c, e, g).
The stimulatory effect of hNmU-25 in human type 2 lymphocytes was confirmed ex-vivo in fresh PBMCs. hNmU-25 enhanced expression of activation markers CD38 and HLA-DR in Th2 and Tc2 cells but not in their CRTH2- counterparts (Fig. 4h, i).
hNmU-25 enhances type 2 cytokine production in human type 2 lymphocytes in response to PGD2 and alarmin stimulationAs we previously reported, mast cell-derived lipid mediators, particularly PGD2 and LTE4, and alarmin proteins (IL-33, IL-25, and TSLP) are important stimulators of type 2 cells in asthma7,24,25,26. We next tested the effect of NmU on cytokine production in human Th2, Tc2, and ILC2 cells in response to these stimulators. Type 2 cytokine production in response to hNmU-25, PGD2, LTE4 and alarmins alone or their combination were compared in these cells (Fig. 5). The efficacy of hNmU-25 on type 2 cytokine production was weaker than that of PGD2, but similar to LTE4 alone (Fig. 5a–c). Synergistic enhancement of IL-5 and IL-13 production by the combination of PGD2 and hNmU-25 was observed in all three cell types except for IL-5 production by Tc2 cells. However, the enhancement of LTE4 stimulation was only observed in IL-5 production by Th2 and ILC2s. The efficacy of hNmU-25 on human type 2 lymphocytes was similar to that of alarmins alone. Moreover, no hNmU-25 effect was observed when hNmU-25 was combined with IL-25, IL-33 or TSLP individually to treat these cells (Fig. 5d–f). However, hNmU-25 additively enhanced IL-5 and IL-13 productions in response to the combination treatment with IL-25/IL-33/TSLP in Th2 and Tc2 cells or with IL-25/33 in ILC2s. These data indicated potential for NmU to enhance type 2 inflammation in asthma.
Fig. 5: NmU enhances stimulatory effects of PGD2 and alarmin proteins in type 2 lymphocytes.
a-c Levels of IL-5 and IL-13 in the supernatants of Th2 (a), Tc2 (b) and ILC2 (c) cultures after treatment with PGD2 or LTE4 alone or in combination with hNmU-25 for 4 h were measured with ELISA. d–f Levels of IL-5 and IL-13 in the supernatants of Th2 (d), Tc2 (e) and ILC2 (f) cultures after treatment with IL-33, IL-25 or TSLP alone or in combination with hNmU-25 for 48 h were determined with ELISA. g, h The levels of mRNA for NMUR1 in cultured Th2 and Tc2 (g) or ILC2 (h) cells after stimulation with hNmU-25 and PGD2 or IL-25/33 measured with qRT-PCR. i NmUR1+ cells detected in cultured Th2 or Tc2 cells with flow cytometry at different time points after treatment with PGD2 or coated anti-CD3/28 antibodies. j NmUR1 expression in ILC2s after treatment with PGD2 (150 nM) or IL-25/33 (50 ng/ml). *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, (n = 7-10 for a, b, d and e; n = 4-5 for c and f; n = 9 for i and j).
To understand the mechanism underlying the synergistic enhancement of hNmU-25 and PGD2, we examined the receptor expression after stimulation of the type 2 cells (Fig. 5g–j; Supplementary Fig. 5). hNmU-25 did not show any effect on the gene transcription of PTGDR2 (encoding CRTH2) (Supplementary Fig. 5a). As expected, PTGDR2 mRNA was slightly downregulated by PGD2 stimulation24. Upregulation of NMUR1 transcription was detected after PGD2 stimulation (Fig. 5g, h). This upregulation was also observed at the protein level after stimulation with PGD2 for 10 h in Th2/Tc2 cells and for 4 h in ILC2s (Fig. 5i, j; Supplementary Fig. 5b). The upregulation of NmUR1 was even stronger in Th2/Tc2 cells via T cell receptor activation or in ILC2s via IL-25/33 stimulation (Fig. 5h–j).
hNmU-25 induces eosinophil migration and degranulationEosinophils play a critical pathogenic role in type 2 immunity-mediated diseases, such as eosinophilic asthma27. It has been reported that human eosinophils expressed NmUR120. We confirmed NmUR1 expression in eosinophils by flow cytometry and Western blot (Fig. 6a, b). Treatment of fresh blood with hNmU-25 ex-vivo induced eosinophil shape change, a marker of eosinophil migration28, in a dose-dependent manner with EC50 = 47.2 nM (Fig. 6c left panel). hNmU-25 (100 nM) also enhanced eosinophil shape change induced by IL-5 (3 ng/ml) although no effect was detected on PGD2-induced eosinophil shape change (Fig. 6c right panel). To confirm the effect of hNmU-25 on eosinophil migration, chemotaxis assay using purified eosinophils was conducted (Fig. 6d). hNmU-25 induced eosinophil migration in a dose-dependent manner peaking at about 100 nM. To investigate the potential impact of NmU on other eosinophil functions, the levels of activation markers, CD11b and CD62L, and degranulation marker, CD63, in eosinophils were measured after treatment with hNmU-25 (Fig. 6e). CD11b and CD62L were unaltered, but CD63 was increased suggesting that NmU could be an inducer of eosinophil degranulation, as evidenced by the morphological changes of eosinophils (Fig. 6f) and the release of eosinophil cationic protein (ECP) after NmU treatment (Fig. 6g).
Fig. 6: NmU induces eosinophil migration and degranulation.
a, b NmUR1 expression in eosinophils was detected with flow cytometry (a) or Western blot (b). c Eosinophil shape-change induced by various concentrations of hNmU-25 or in combination with PGD2 or IL-5 measured with flow cytometry. d Eosinophil migration induced by various concentrations of hNmU-25 determined with chemotaxis assay. e Surface levels of CD11b, CD62L, and CD63 in eosinophils after treatment with hNmU-25 measured with flow cytometry. f Representative image of eosinophil degranulation induced by hNmU-25. g Levels of ECP in the supernatants of eosinophils after treatment with hNmU-25 for 4 h were measured with ELISA. IL-5 stimulation was used as a positive control. *p < 0.05, **p < 0.01, ***p < 0.001, ****P < 0.0001, (n = 3–5 for c; n = 4 for d; n = 5 for g).
留言 (0)