
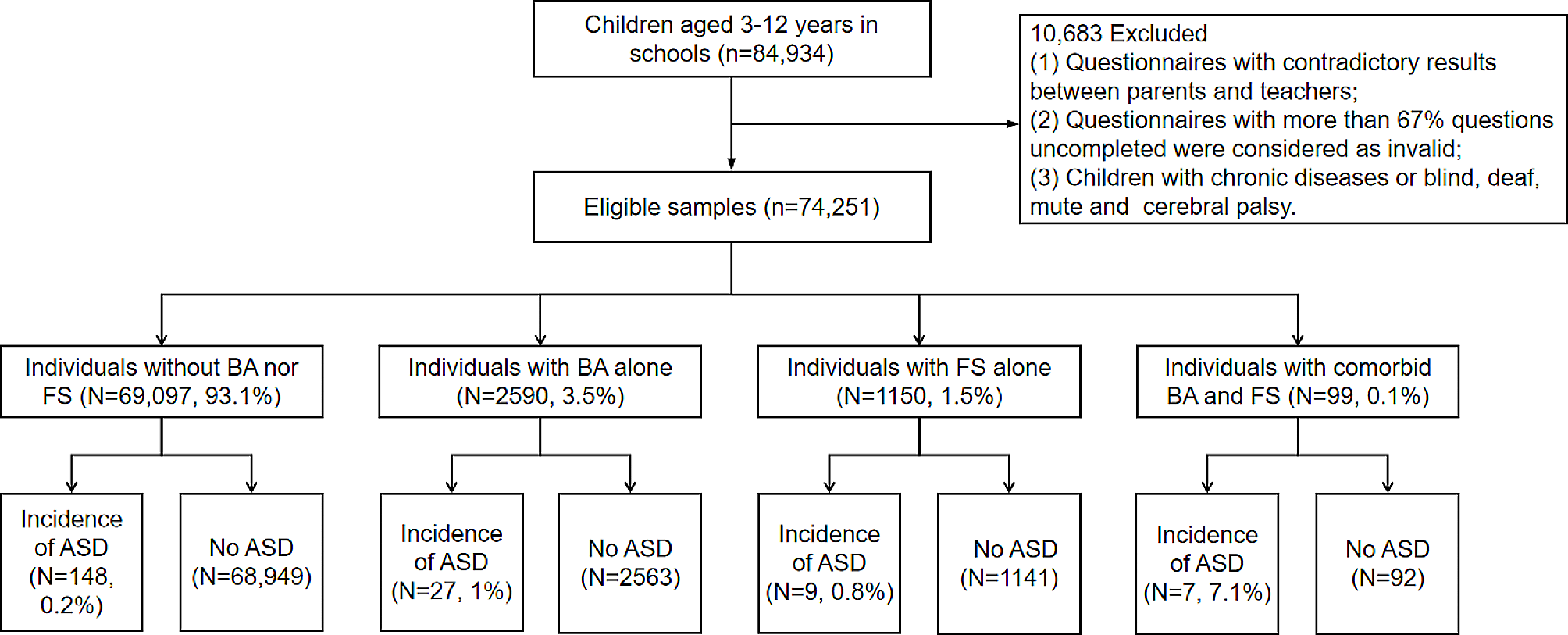
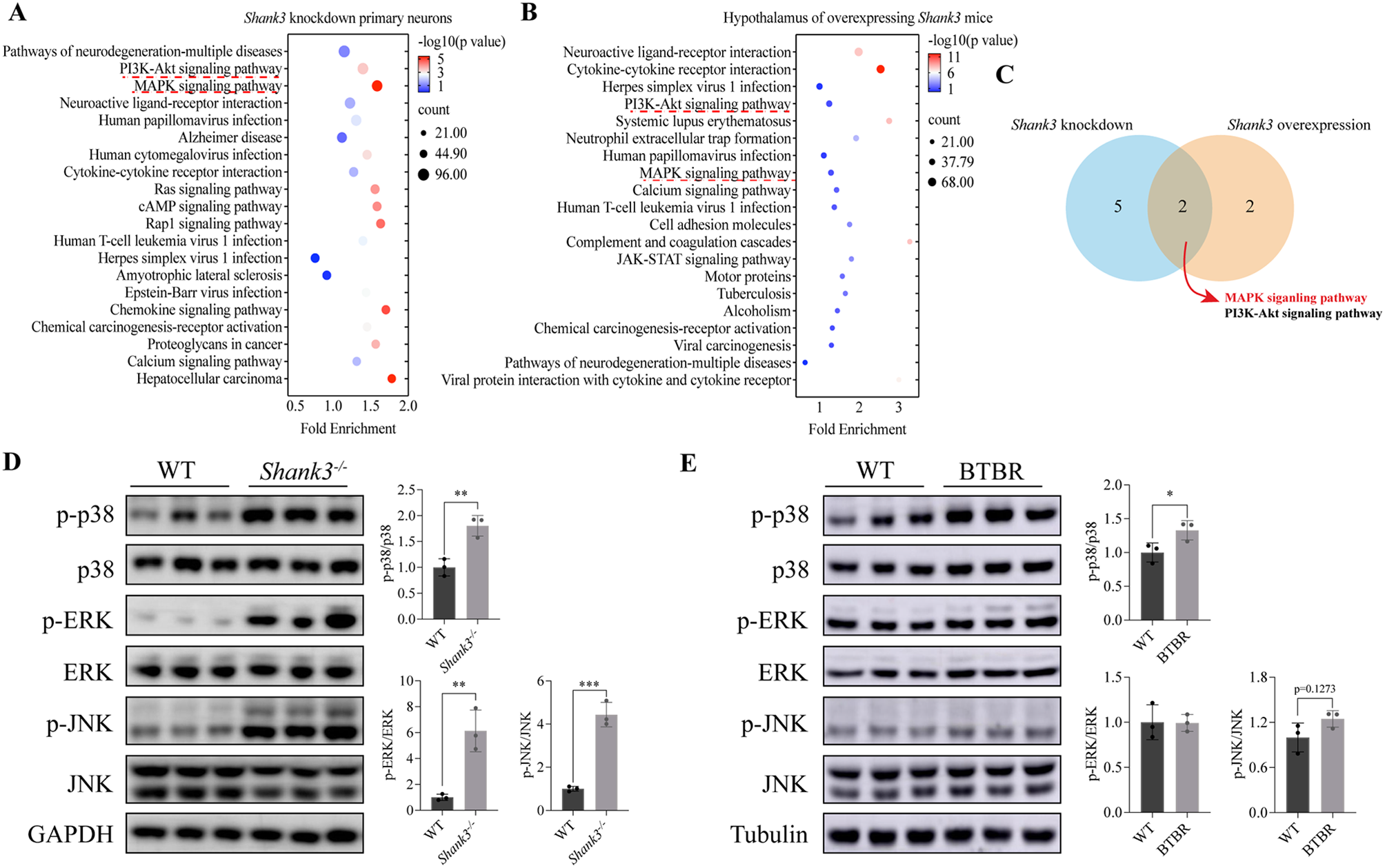
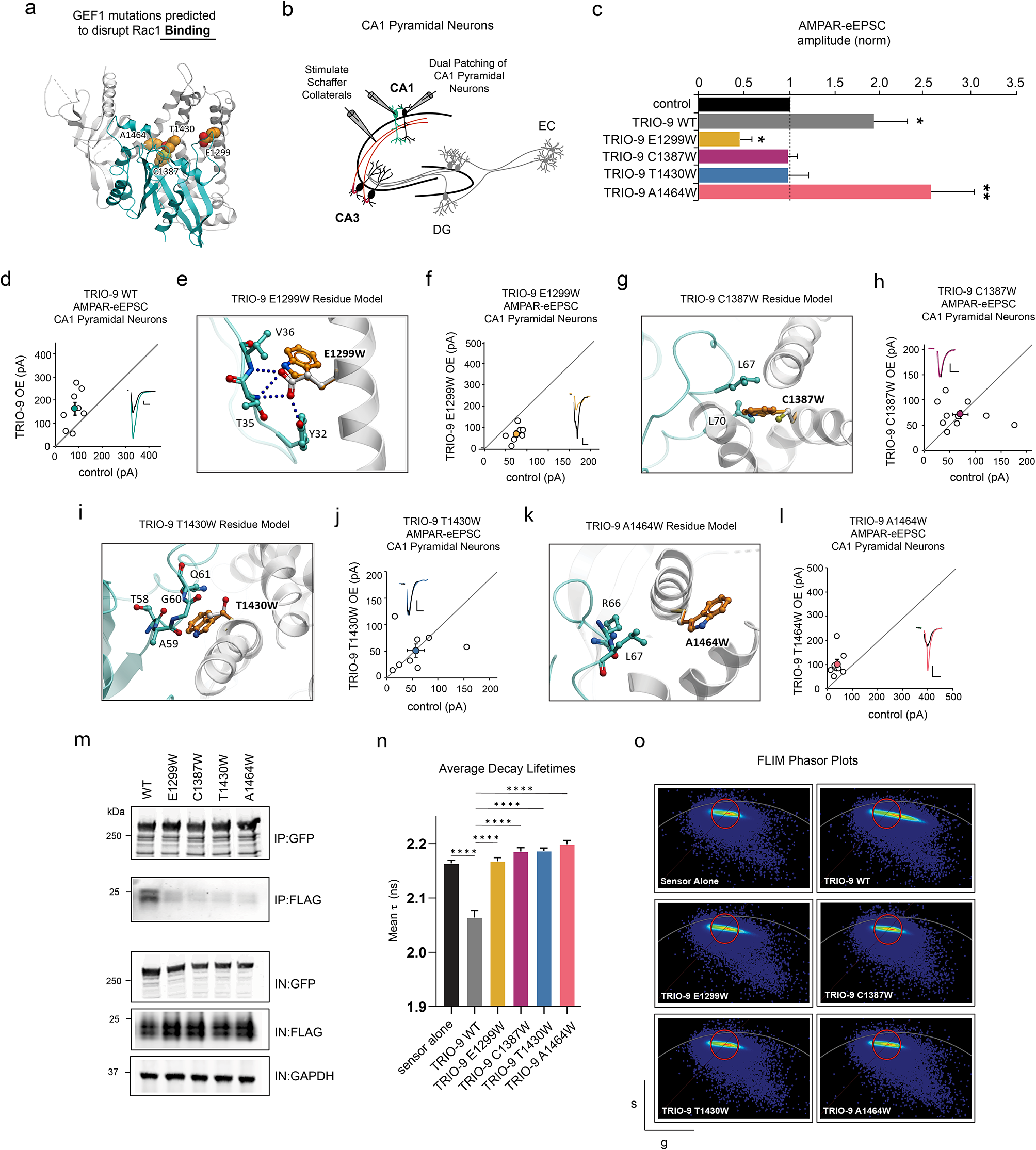
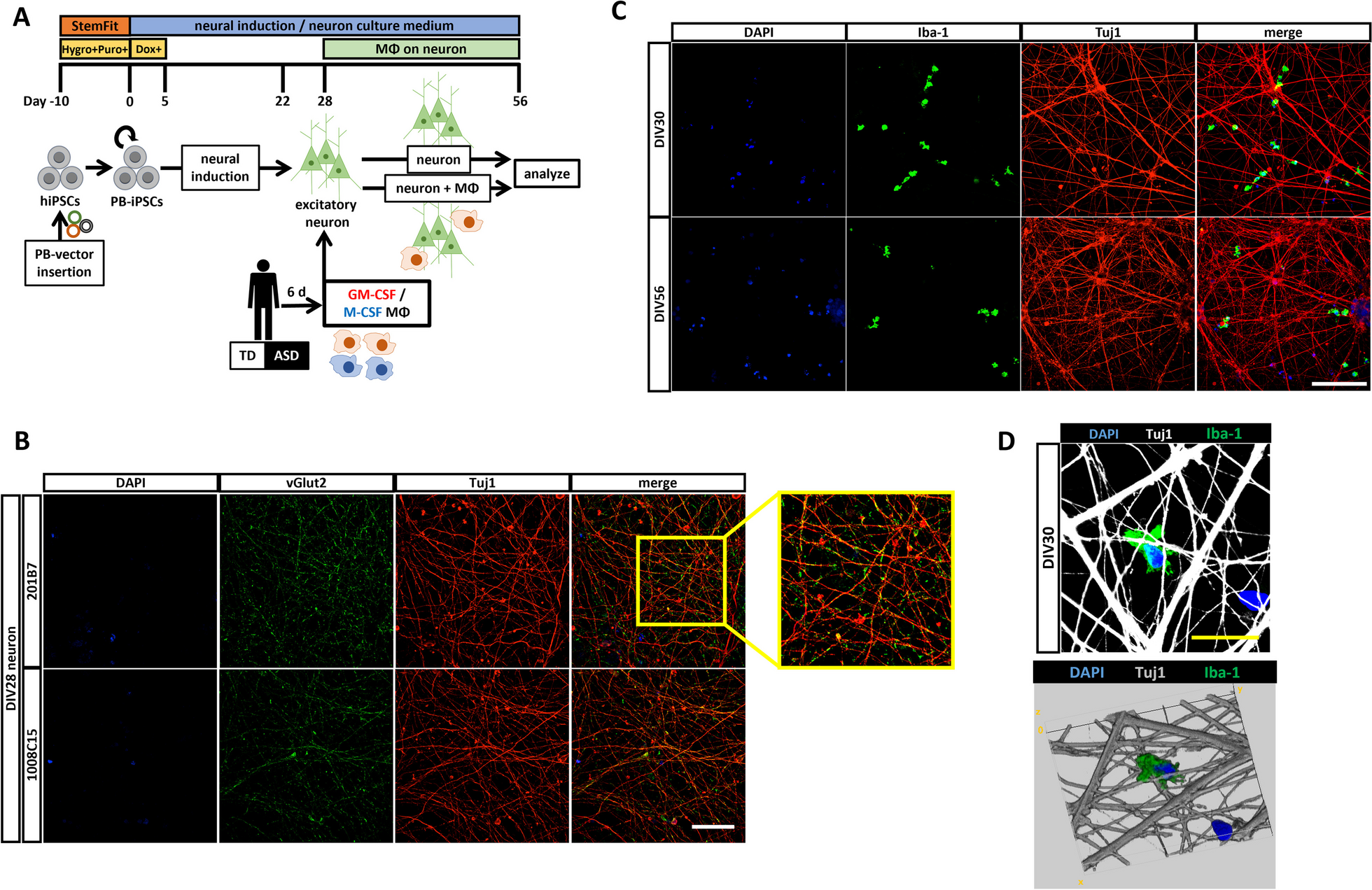
記住我






Elevated de novo protein synthesis is one of the core pathophysiological phenotypes reported in Fmr1−/y mouse brain [11]. However, assessment of de novo protein synthesis in specific populations of inhibitory neurons has not been carried out in Fmr1−/y mice [12]. To explore the role of FMRP in PV and SOM-positive inhibitory neurons, we first examined FMRP expression in these neuronal populations in the mouse hippocampus. We performed immunostaining for FMRP and PV or SOM in wild type (WT) mouse hippocampus and found that FMRP signal localized to both PV and SOM-expressing cells, whereas FMRP labeling was absent in these cells in the Fmr1−/y mouse hippocampus (Additional file 1: Fig. S1). Next, to quantify the levels of de novo protein synthesis in PV and SOM-expressing neurons, we performed fluorescent non-canonical amino acid tagging (FUNCAT) along with immunostaining for PV and SOM in hippocampal slices from Fmr1−/y and WT littermates (Fig. 1A, B). The levels of PV and SOM expression were not changed in Fmr1−/y compared to WT mouse hippocampus (Fig. 1C , p > 0.1 and Fig. 1E, p = 0.059, Student’s t test, NS). However, the FUNCAT signal was significantly elevated in Fmr1−/y mouse PV and SOM-positive neurons in the hippocampus compared to WT (Fig. 1D, ***p < 0.0001 and Fig. 1F, ***p < 0.0001, Student’s t test) and was near background levels in Fmr1−/y and WT hippocampal slices where azidohomoalanine (AHA) was omitted (Fig. 1G). Thus, consistent with previous studies of several brain regions of the Fmr1−/y mouse [11, 12, 27], de novo protein synthesis was increased in PV and SOM-positive neurons in the hippocampus of Fmr1−/y mice.
Fig. 1
De novo protein synthesis is elevated in PV and SOM-positive neurons in Fmr1−/y mouse hippocampus. A–B Representative images of FUNCAT in hippocampal slices from WT and Fmr1−/y mice, co-stained for either PV A or SOM (B, scale bar: 50 μm). C–D Quantification of PV labeling C or AHA fluorescence D in PV-positive WT and Fmr1−/y mouse hippocampal cells. Values represent mean ± SEM (n = 20–25 z-stacks, 9–11 sections, from 3 animals per genotype). E–F Quantification of SOM labeling E or AHA fluorescence F in SOM-positive WT and Fmr1−/y mouse hippocampal cells. Values represent mean ± SEM (n = 26–39 z-stacks, 11–13 sections from 3 animals per genotype). G Representative images of FUNCAT control in WT and Fmr1−./y mouse hippocampal slices where AHA was omitted; ***p < 0.0001 (Student’s t test)
Mice with cell-type specific deletion of Fmr1 in PV-positive neurons exhibit mild anxiety-like behaviors and impaired social interactionTo determine the cell type-specific contribution of FMRP expression in PV-positive neurons to autistic-like behavioral deficits, we utilized Cre-lox technology to generate mice with a cell type-specific deletion of Fmr1 in PV-expressing neurons, termed Fmr1flx/flx:Pv-Cre (Fmr1−/y-PV) mice. First, we confirmed the lack of FMRP expression in PV-positive neurons in Fmr1−/y-PV mouse mPFC (Fig. 2A). We found that approximately 49% and 59% of PV-positive cells expressed FMRP in WT-PV mouse mPFC and hippocampus, respectively. We quantified FMRP fluorescence in all PV-expressing cells in WT-PV and Fmr1−/y-PV mice and observed a significant reduction of FMRP expression in Fmr1−/y-PV mPFC and hippocampus (Additional file 1: Fig. S2A, ***p < 0.0001 and B, ***p < 0.0001, Student’s t test). We believe the residual fluorescent signal observed in Fmr1−/y-PV was due to nonspecific FMRP antibody signal in FMRP negative cells (see staining in Fmr1−/y mice, Additional file 1: Fig. S1, bottom panel). Next, we performed a battery of tests to assess motor function, anxiety, repetitive, stereotypic, and social behaviors in Fmr1+/+:Pv-Cre (WT-PV) and Fmr1−/y-PV mice. Fmr1−/y-PV mice displayed normal motor behavior and learning (Additional file 1: Fig. S2C, two-way RM ANOVA, followed by Bonferroni’s multiple comparisons test; trial x genotype, Day 1, F(3, 72) = 0.599, p > 0.1; Day 2, F(3, 72) = 1.552, p > 0.1). Using elevated plus maze (EPM), we assessed anxiety-like behavior in WT-PV and Fmr1−/y-PV mice. Time spent in the open arms of EPM was not different between the genotypes (mean ± SEM: WT-PV 41.81 s ± 10.71, Fmr1−/y-PV 28.04 s ± 6.75, p > 0.1, Student’s t test), but Fmr1−/y-PV mice spent significantly more time than WT-PV mice in the closed arms (mean ± SEM: WT-PV 161.5 s ± 11.98, Fmr1−/y-PV 197.3 s ± 11.40, *p = 0.041, Student’s t test) but significantly less time in the center (mean ± SEM: WT-PV 90.60 s ± 7.05, Fmr1−/y-PV 69.48 s ± 6.38, *p = 0.036, Student’s t test) of the EPM (Fig. 2B), indicative of elevated anxiety.
Fig. 2
Mice lacking Fmr1 in PV-expressing neurons exhibit mild anxiety-like behaviors and impaired social interaction. A Representative images of mPFC sections from WT-PV and Fmr1−/y-PV mice, immunostained for PV and FMRP (scale bar: 50 μm). Bottom panels show enlarged boxed regions of individual PV-positive cells. B Time spent in the open arms of EPM was not significantly different between the genotypes (left), whereas Fmr1−/y-PV mice spent more time in the closed arms (middle) and less time in the center (left) of the EPM; mean ± SEM (n = 12–14 animals per genotype). C Time spent self-grooming and D total number of marbles buried, as well as time spent burying/digging up marbles did not differ significantly between WT-PV and Fmr1−/y-PV mice. Values represent mean ± SEM (n = 10–14 animals per genotype). E Fmr1−/y-PV mice made significantly greater percentage of errors during the training portion of the Y Maze task, whereas their performance during LTM test and reversal portions was not significantly different between the genotypes. Values represent mean ± SEM (n = 12–13 animals per genotype). F Fmr1−/y-PV and WT-PV mice spent more time sniffing a stranger mouse (S) compared to an object (O) during Sociability portion of the 3CSI Test. Values represent mean ± SEM (n = 12–14 animals per genotype). Preference Index (PI) for a stranger mouse was not different between the genotypes. G WT-PV mice spent significantly more time sniffing a novel stranger mouse (S2) compared to a familiar stranger mouse (S1) during the Social Novelty test, while Fmr1−./y-PV mice exhibited impaired social novelty behavior. (n = 12–14 animals per genotype). PI for a S2 was also significantly different between the genotypes; *p < 0.05; **p < 0.01; ***p < 0.001; Two-way ANOVA or RM two-way ANOVA followed by Bonferroni’s multiple comparisons test, Student’s t test
We used self-grooming and marble bury tests to assess if Fmr1−/y-PV mice exhibit repetitive and stereotypic behaviors, one of the criteria for ASD diagnosis [28]. Time spent self-grooming (mean ± SEM: WT-PV 60.15 s ± 8.75, Fmr1−/y-PV 66.77 s ± 10.24, p > 0.1, Student’s t test), the total number of marbles buried (mean ± SEM: WT-PV 13.67 ± 1.38, Fmr1−/y-PV 14.07 ± 1.57, p > 0.1, Student’s t test), or time spent burying/digging up marbles (two-way RM ANOVA, followed by Bonferroni’s multiple comparisons test; time x genotype, F(2, 42) = 1.389, p > 0.1) was not significantly different between the genotypes (Fig. 2C, D). Thus, Fmr1−/y-PV mice do not exhibit these forms of repetitive behaviors. Rigidity in behavior and the need for sameness are manifestations of perseverative behaviors associated with ASD [28]. We used a water-based Y-maze task to assess cognitive flexibility in mice with cell type-specific deletion of Fmr1. Fmr1−/y-PV mice were impaired during initial acquisition portion of the task as evident by a significant decrease in percentage of correct arm choices during training (Fig. 2E, training portion, two-way RM ANOVA, followed by Bonferroni’s multiple comparisons test; time x genotype, F(3, 69) = 1.977, p > 0.1; a significant difference between genotypes during trial block 2 using post-hoc Bonferroni’s multiple comparisons test, *p = 0.027). However, after a criterion level of performance was achieved, Fmr1−/y-PV mice performed similar to WT-PV mice during long-term memory (LTM) testing and reversal portions of the test (Fig. 2E, LTM, p > 0.1, Student’s t test, NS; reversal, two-way RM ANOVA, followed by Bonferroni’s multiple comparisons test; time x genotype, F(6, 138) = 0.583, p > 0.1). Therefore, mice with Fmr1 deletion in PV neurons exhibit normal cognitive flexibility.
Deficits in social interaction represent another group of diagnostic criteria associated with ASD [28]. In the sociability portion of the 3-chamber social interaction (3CSI) test, both Fmr1−/y-PV and WT-PV mice spent more time exploring and sniffing a novel mouse (S) compared to an object (O), Fig. 2F, two-way ANOVA, followed by Bonferroni’s multiple comparisons test, F(1, 48) = 64.74, ***p < 0.0001). In addition, there was no significant difference between the genotypes in the preference index (PI) for the novel mouse, indicating normal social preference behavior in Fmr1−/y-PV mice (Fig. 2F, p > 0.1, Student’s t test). On the other hand, during the social novelty portion of the test, WT-PV mice displayed a natural preference to spend more time exploring/sniffing a novel mouse (stranger 2, S2), whereas Fmr1−/y-PV did not show a preference for interacting with S2 (Fig. 2G, two-way ANOVA, followed by Bonferroni’s multiple comparisons test, F(1, 48) = 11.34, **p < 0.01, p > 0.1). Furthermore, Fmr1−/y-PV had a significantly decreased PI for S2, suggesting they did not exhibit a preference for social novelty (Fig. 2G, p = 0.041, Student’s t test). This was unlikely due to their mild anxiety, as the total time spent interacting with S1 + S2 during social novelty test did not differ between the genotypes (mean total interaction time ± SEM: WT-PV 116.8 s ± 9.98, Fmr1−/y-PV 119.8 s ± 10.22, p = 0.831, Student’s t test). Taken together, these findings suggest that ablation of FMRP expression in PV-positive neurons results in a specific pattern of ASD-like behavioral deficits (Table 1).
Table 1 Summary of behavioral alterations in Fmr1−/y-PV and Fmr1−./y-SOM mice compared to control Fmr1 −/y-PV mice do not display deficits in learning and memoryIntellectual disability is a prominent feature of FXS affecting over 85% of males and 25% of females to varying levels of severity [4, 5]. To examine the impact of Fmr1 deletion in PV positive neurons on learning and memory, we carried out a series of cognitive tasks with the Fmr1−/y-PV and WT mice. During the object recognition task, Fmr1−/y-PV and WT-PV mice showed a preference for the novel object during both short-term (STM) (two-way ANOVA, followed by Bonferroni’s multiple comparisons test, F(1, 46) = 324.2, ***p < 0.0001) and long-term memory (LTM) (two-way ANOVA, followed by Bonferroni’s multiple comparisons test, F(1, 46) = 102.0, ***p < 0.0001) portions of the test (Fig. 3A, B). In addition, PI for novel object during STM and LTM was not different, indicating intact memory formation and recall (Additional file 1: Fig. S3A, p > 0.1, Student’s t test).
Fig. 3
Mice with Fmr1 deletion in PV-positive neurons do not display deficits in learning and memory. A, B Fmr1−/y-PV and WT-PV mice spent more time exploring a novel object during STM A and LTM B portions of the object recognition test. Values represent mean ± SEM (n = 12–13 animals per genotype). C Escape latency during the training portion of MWM was not significantly different between Fmr1−/y-PV and WT-PV mice. Values represent mean ± SEM (n = 15 animals per genotype). D, E Target quadrant occupancy D or frequency of platform crossings during 60 s probe trial of MWM test were not significantly different between Fmr1−/y-PV and WT-PV mice. Values represent mean ± SEM (n = 15 animals per genotype). F Preference index for a novel object location during OLM test was not significantly different between Fmr1−/y-PV and WT-PV mice. Values represent mean ± SEM (n = 8–10 animals per genotype). G, H Following threat conditioning, Fmr1−/y-PV mice spent similar percentage of time freezing to the context G but spent significantly more time freezing to the audible cue H compared to WT-PV mice. Values represent mean ± SEM (n = 12–14 animals per genotype). I Acquisition of threat memory extinction was not significantly different between the genotypes. Freezing to the tone shown in trial blocks on day 1 and day 2 of threat memory extinction. Values represent mean ± SEM (n = 12–14 animals per genotype). J Fmr1−/y-PV mice spent significantly more time freezing to the audible cue following reactivation of auditory threat memory. Values represent mean ± SEM (n = 12–14 animals per genotype); ***p < 0.001; *p < 0.05; Two-way ANOVA or RM two-way ANOVA followed by Bonferroni’s multiple comparisons test, Student’s t test
We next tested spatial memory in Fmr1−/y-PV and WT mice using Morris Water Maze (MWM). Escape latency during training did not differ significantly between the genotypes (Fig. 3C, two-way RM ANOVA, followed by Bonferroni’s multiple comparisons test; time x genotype, F(3, 84) = 0.017, p > 0.1). On the probe test day, Fmr1−/y-PV mice spent more time in the target quadrant of the maze, similar to WT-PV mice (Fig. 3D, two-way RM ANOVA, followed by Bonferroni’s multiple comparisons test, quadrant x genotype, F(3, 84) = 0.674, p > 0.1). Even though Fmr1−/y-PV mice showed a trend for reduced platform crossings during the probe test, it did not reach significance (Fig. 3E, p > 0.1, Student’s t test). In addition, escape latency during MWM reversal portion was not significantly different between the genotypes (Additional file 1: Fig. S3B, two-way RM ANOVA, followed by Bonferroni’s multiple comparisons test, time x genotype, F(2, 56) = 1.473, p > 0.1). Fmr1−/y-PV mice did not exhibit vision impairments that could affect their performance during MWM, as there was no difference in escape latency between the genotypes when the platform was marked by a visible cue (Additional file 1: Fig. S3C, p > 0.1, Student’s t test). To further discern whether Fmr1−/y-PV mice exhibit deficits in hippocampus-dependent spatial memory, we used the object location memory (OLM) task. PI for the object in a novel versus familiar location did not differ between Fmr1−/y-PV and WT-PV mice, indicating intact hippocampal memory (Fig. 3F, p > 0.1, Student’s t test). Lastly, we performed a threat conditioning task to test the associative memory of Fmr1−/y-PV and WT-PV mice. Fmr1−/y-PV mice spent similar percentage of time freezing to the context associated with the shocks 24 h after training (Fig. 3G, p > 0.1, Student’s t test) but spent significantly more time freezing during tone presentation compared to WT-PV mice (Fig. 3H, p = 0.014, Student’s t test). Acquisition of threat memory extinction was not significantly different between the genotypes (Fig. 3I, two-way RM ANOVA, followed by Bonferroni’s multiple comparisons test, time block x genotype, Day 1, F(4, 96) = 1.849, p > 0.1; Day 2, F(4, 96) = 1.050, p > 0.1). In addition, Fmr1−/y-PV mice spent significantly more time freezing during tone presentation compared to WT-PV during reconsolidation portion of threat conditioning test (Fig. 3J, p = 0.018, Student’s t test). Together, these findings indicate that absence of FMRP in PV positive neurons is not associated with any significant deficits in learning and memory but may play a role in limiting long-term auditory threat memory and memory reconsolidation (Table 1).
Deletion of Fmr1 in SOM-positive neurons does not result in autistic-like or cognitive behavioral deficitsAs PV- and SOM-positive inhibitory neurons play different roles in circuit function and behavior, we wanted to dissect the cell type-specific role of FMRP in SOM-expressing neurons in autism-associated behaviors and cognition. To this end, we generated mice with Fmr1 deletion in SOM-expressing neurons, the Fmr1flx/flx: Sst-IRES-Cre (Fmr1−/y-SOM mice) and performed behavioral testing with the Fmr1−/y-SOM and Fmr1+/+: Sst-IRES-Cre (WT-SOM) mice. First, the lack of FMRP expression in SOM-positive inhibitory neurons was confirmed in Fmr1−/y-SOM mouse mPFC (Fig. 4A). Approximately 67% and 50% of SOM-positive cells expressed FMRP in WT-SOM mouse mPFC and hippocampus, respectively. FMRP signal was quantified in all SOM-expressing cells in WT-SOM and Fmr1−/y-SOM mice and a significant reduction of FMRP expression was observed in Fmr1−/y-SOM mPFC and hippocampus (Additional file 1: Fig. S4A, ***p < 0.0001 and 4B, ***p = 0.0001, Student’s t test). Once again, there was residual fluorescent signal observed in Fmr1−/y-PV due to nonspecific FMRP antibody signal in FMRP negative cells (see staining in in Fmr1−/y mice, Additional file 1: Fig. S1, bottom panel). We proceeded to perform behavioral experiments and observed no difference in the amount of time spent in the open or closed arms of the EPM between WT-SOM and Fmr1−/y-SOM mice (Fig. 4B, time spent in the open arm, mean ± SEM: WT-SOM 42.77 s ± 9.23, Fmr1−/y-SOM 36.81 s ± 7.64, p > 0.1; time spent in the closed arm, mean ± SEM: WT-SOM 182.6 s ± 14.08, Fmr1−/y-SOM 195.4 s ± 12.16, p > 0.1, Student’s t test), indicating that Fmr1−/y-SOM mice do not exhibit anxiety-like behavior. In addition, Fmr1−/y-SOM mice did not exhibit repetitive, stereotypic behaviors as the number of marbles buried (mean ± SEM: WT-SOM 9.11 ± 2.00, Fmr1−/y-SOM 7.25 ± 1.59, p > 0.1, Student’s t test) and the time spent self-grooming (mean ± SEM: WT-SOM 48.52 s ± 7.43, Fmr1−/y-SOM 60.15 s ± 8.83, p > 0.1, Student’s t test) were not significantly altered compared to WT-SOM mice (Fig. 4C, D, respectively). In addition, we did not observe perseverative behavior in Fmr1−/y-SOM mice using the water-based Y-maze as percentage of correct arm choices was not different during training, LTM test or reversal portions of the test (Additional file 1: Fig. S4C, training portion, two-way RM ANOVA, followed by Bonferroni’s multiple comparisons test; time x genotype, F(3, 57) = 1.409, p > 0.1; LTM, p > 0.1, Student’s t test, NS; reversal portion, two-way RM ANOVA, followed by Bonferroni’s multiple comparisons test; time x genotype, F(6, 114) = 0.744, p > 0.1). Overall, these data indicate that Fmr1−/y-SOM mice did not exhibit anxiety-like, stereotypic, or perseverative behaviors.
Fig. 4
Fmr1 deletion in SOM-expressing neurons does not result in cognitive or autistic-like deficits in mice. A Representative images of mPFC sections from WT-SOM and Fmr1−/y-SOM mice, immunostained for SOM and FMRP (scale bar: 50 μm). Bottom panels show enlarged boxed regions of individual SOM-positive cells in WT-SOM and Fmr1−/y-SOM mouse mPFC. B Time spent in the open or closed arms of the EPM, C number of marbles buried or D time spent self-grooming were not significantly different between WT-SOM and Fmr1−/y-SOM mice. Values represent mean ± SEM (n = 9–12 animals per genotype). E Preference Index (PI) for a stranger mouse (S) during Social Preference or a novel stranger mouse (S2) during Social Novelty portion were not significantly different between the genotypes. Values represent mean ± SEM (n = 9–12 animals per genotype). F Fmr1−/y-SOM and WT-SOM mice spent more time exploring a novel object during STM and LTM portions of the object recognition test. Values represent mean ± SEM (n = 9–12 animals per genotype). G, H Escape latency during the training portion of MWM G or frequency of platform crossings during 60 s probe trial H were not significantly different between Fmr1−/y-SOM and WT-SOM mice. Values represent mean ± SEM (n = 9–12 animals per genotype). I–K Fmr1−./y-SOM mice spent similar percentage of time compared to WT-SOM mice in freezing behavior in the context associated with the shocks (I, contextual threat memory), as well as during tone presentation (J, cued threat memory). In addition, there was no significant difference between the genotypes in time spent freezing to the audible cue following reactivation of auditory threat memory (K). Values represent mean ± SEM (n = 9–12 animals per genotype); ***p < 0.001; *p < 0.05; Two-way ANOVA or RM two-way ANOVA followed by Bonferroni’s multiple comparisons test, Student’s t test
In the 3CSI test, both WT-SOM and Fmr1−/y-SOM mice spent longer time sniffing and exploring novel S mouse over an object (Additional file 1: Fig. S4D, two-way ANOVA, followed by Bonferroni’s multiple comparisons test, F(1, 38) = 36.19, ***p < 0.0001), and PI for S was not different between the genotypes (Fig. 4E, p > 0.1, Student’s t test), indicating that Fmr1−/y-SOM mice exhibit normal social preference behavior. In the social novelty portion of the 3CSI, there was an overall trend for WT-SOM and Fmr1−/y-SOM mice to spend more time sniffing S2, however it was not significant (Additional file 1: Fig. S4E, two-way ANOVA, followed by Bonferroni’s multiple comparisons test, F(1, 38) = 2.901, p > 0.1). In addition, PI for S2 was at or below 0.5 (no preference) for 3/9 of WT-SOM mice and 6/12 of Fmr1−/y-SOM mice (Fig. 4E, p > 0.1, Student’s t test). Thus, overall social behavior was unaffected in Fmr1−/y-SOM mice compared to WT-SOM, but both genotypes displayed decreased preference for social novelty.
Assessment of learning and memory was performed in WT-SOM and Fmr1−/y-SOM mice. During the object recognition task, WT-SOM and Fmr1−/y-SOM mice spent longer time exploring a novel compared to familiar object during STM and LTM (Fig. 4F, STM two-way ANOVA, followed by Bonferroni’s multiple comparisons test, F(1, 19) = 48.12, ***p < 0.0001, **p < 0.001; LTM two-way ANOVA, followed by Bonferroni’s multiple comparisons test, F(1, 38) = 135.8, ***p < 0.0001). PI for novel object was also unchanged in Fmr1−/y-SOM mice (Additional file 1: Fig. S4F and G, p > 0.1, Student’s t test). Spatial memory in MWM was unaltered in Fmr1−/y-SOM mice; escape latency during training was similar between WT-SOM and Fmr1−/y-SOM mice (Fig. 4G, two-way RM ANOVA, followed by Bonferroni’s multiple comparisons test; time x genotype, F(3, 57) = 0.622, p > 0.1), as well as time spent in the target quadrant (Additional file 1: Fig. S4H, two-way RM ANOVA, followed by Bonferroni’s multiple comparisons test, quadrant x genotype, F(3, 57) = 0.086, p > 0.1) and number of platform crossings (p > 0.1, Student’s t test) on probe test day (Fig. 4H). Escape latency during MWM reversal (two-way RM ANOVA, followed by Bonferroni’s multiple comparisons test, time x genotype, F(2, 38) = 0.017, p > 0.1) and visible platform test (p > 0.1, Student’s t test) also did not differ between the genotypes (Additional file 1: Fig. S4I, J). Lastly, Fmr1−/y-SOM mice were tested in threat conditioning task and percentage of time spent in freezing behavior in the context associated with shocks and during tone presentation (Fig. 4I, J, respectively, p > 0.1, Student’s t test) or during reactivation of auditory threat memory were not different from WT-SOM (Fig. 4K, p > 0.1, Student’s t test). Overall, we did not observe significant autism-related or cognitive deficits in Fmr1−/y-SOM mice (Table 1).
Protein synthesis is dysregulated in PV, but not SOM-positive neurons following cell type-specific deletion of Fmr1To elucidate molecular alterations arising from cell type-specific ablation of FMRP, we examined PV and SOM expression and de novo protein synthesis using immunostaining and FUNCAT in mPFC and hippocampus (Fig. 5A, B), forebrain regions relevant to the behaviors examined in Figs. 2, 3, and 4. We found that expression of PV itself was elevated both in Fmr1−/y-PV hippocampus and mPFC compared to WT-PV (Fig. 5C, hippocampus **p < 0.01; mPFC; *p < 0.05, Student’s t test). Furthermore, we found that de novo protein synthesis was elevated in PV-positive neurons in Fmr1−/y-PV mPFC but decreased in the hippocampus (Fig. 5D, E, respectively, hippocampus ***p < 0.001; mPFC ***p < 0.001, Student’s t test). To further characterize brain region-specific cellular deficits in regulation of protein synthesis by FMRP in PV-expressing neurons, we examined phosphorylation of ribosomal protein S6 (rpS6), a downstream effector of mammalian target of rapamycin complex 1 (mTORC1), in Fmr1−/y-PV mPFC and hippocampus. Signaling by mTORC1 leads to activation of protein synthesis and both processes are elevated in Fmr1−/y mice [27, 29]. Importantly, phosphorylation of rpS6 was also enhanced in Fmr1−/y mouse hippocampal neurons [15]. Our results indicate that rpS6 phosphorylation (Ser 235/6) was increased in PV-positive hippocampal neurons but unaltered in PV-positive inhibitory neurons in mPFC of Fmr1−/y-PV mice (Additional file 1: Fig. S5, hippocampus *p < 0.016; mPFC p > 0.1, Student’s t test). Thus, there is a brain region-specific dysregulation of de novo protein synthesis and mTORC1 downstream signaling upon cell type-specific deletion of Fmr1 in PV-positive inhibitory neurons, which may underlie the specific behavioral deficits (Additional file 2: Table S1).
Fig. 5
Brain region-specific cellular deficits in PV- and SOM-positive neurons upon cell type-specific deletion of Fmr1. A, B Representative images of FUNCAT and PV staining from WT-PV and Fmr1−/y-PV mouse mPFC (A, scale bar: 50 μm) or FUNCAT and SOM staining from WT-SOM and Fmr1−/y-SOM mice (B). Arrows denote individual PV- or SOM-expressing cells with FUNCAT signal. C PV expression was elevated in mPFC and hippocampus of Fmr1−/y-PV mouse. Quantification of PV labeling in PV-positive neurons in WT-PV and Fmr1−/y-PV mouse mPFC and hippocampus. D, E Cell type-specific deletion of Fmr1 results in elevated de novo protein synthesis in PV-positive neurons in mPFC, but a decrease in hippocampus of Fmr1−/y-PV mice. Quantification of AHA fluorescence in PV-positive neurons in WT-PV and Fmr1−/y-PV mouse mPFC D and hippocampus E. Values represent mean ± SEM (hippocampus, n = 45–48 z-stacks, 19–24 sections, from 3 animals per genotype; mPFC, n = 24–25 z-stacks, 14 sections, from 3 animals per genotype); ***p < 0.0001 (Student’s t test). F SOM expression was elevated in mPFC but decreased in hippocampus of Fmr1−/y-SOM mice. Quantification of SOM labeling in SOM-positive neurons in WT-SOM and Fmr1−/y-SOM mouse mPFC and hippocampus. G, H De novo protein synthesis is not significantly different upon cell type-specific deletion of Fmr1 in SOM-positive neurons. Quantification of AHA fluorescence in SOM-positive neurons in WT-SOM and Fmr1−./y-SOM mouse mPFC G and hippocampus (H). Values represent mean ± SEM (hippocampus, n = 26–34 z-stacks, 11–14 sections, from 2 animals per genotype; mPFC, n = 39 z-stacks, 21–22 sections, from 3 animals per genotype); ***p < 0.0001 (Student’s t test)
Expression of SOM was significantly increased in mPFC but decreased in the hippocampus of Fmr1−/y-SOM (Fig. 5F, hippocampus *** p < 0.001; mPFC ***p < 0.001, Student’s t test). On the other hand, de novo protein synthesis in SOM-positive inhibitory neurons did not differ between WT-SOM and Fmr1−/y-SOM mice, both in mPFC and hippocampus (Fig. 5G, H, respectively, hippocampus p > 0.1; mPFC; p > 0.1, Student’s t test). Similar to findings from behavioral experiments, global de novo protein synthesis was unaffected by cell type-specific deletion of Fmr1 in SOM-expressing inhibitory neurons. Together, these findings indicate that PV neurons appear to be particularly sensitive to FMRP ablation at the molecular and behavioral level (Additional file 2: Table S1).
留言 (0)