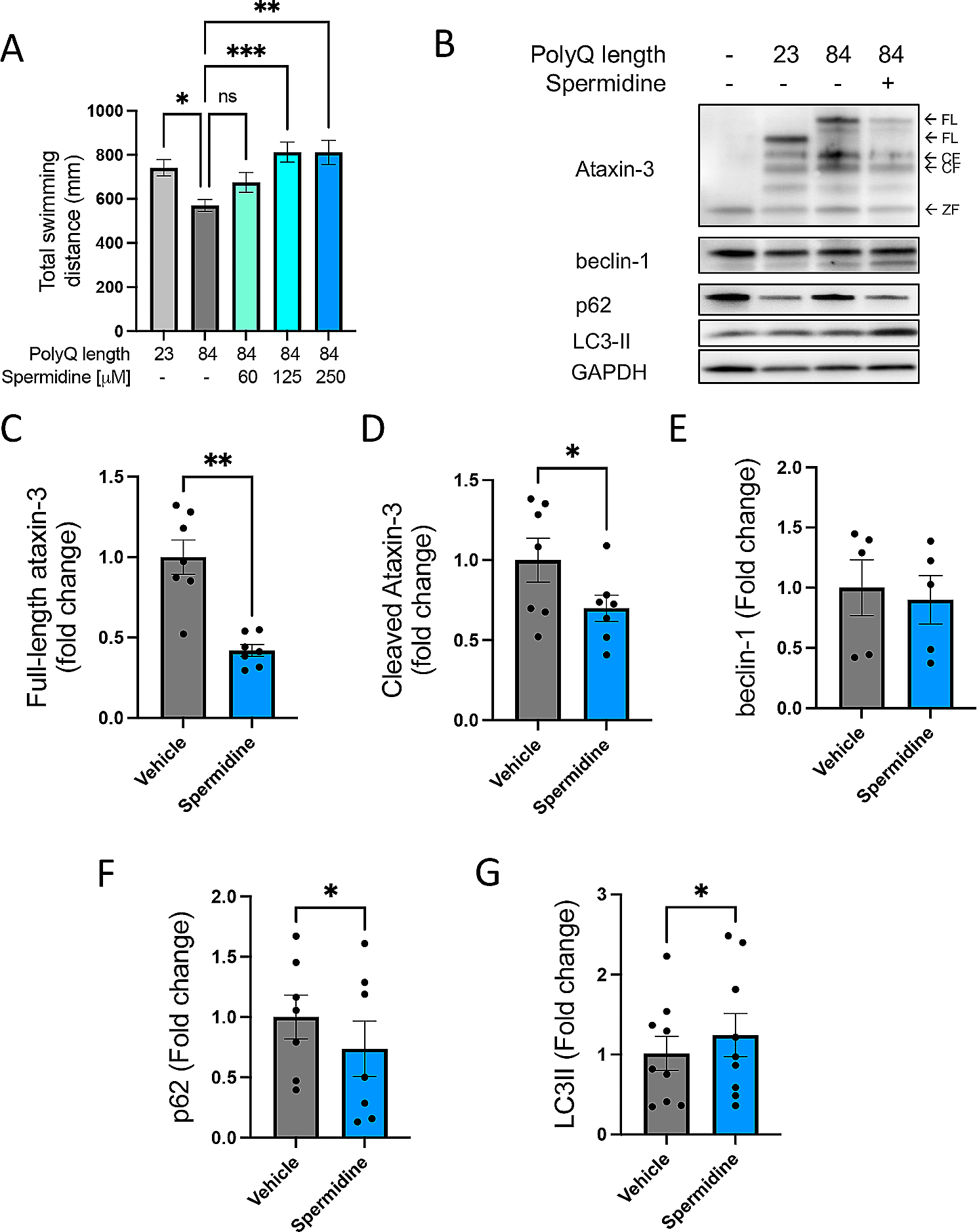
In this study, the concentrations of GFAP, MBP, NFL, tau and UCHL1 were determined in 17 anatomical regions of the CNS. Our findings indicate a substantial regional variation in the concentration of the investigated proteins. The concentration of GFAP was twenty times higher in the medulla oblongata and cervical spinal cord, compared with cerebral cortex. The concentration of MBP in highly myelinated areas of CNS (cerebral white matter, pons, medulla oblongata and cervical spinal cord) was tenfold higher than in cerebral cortex. In contrast, tau had an inverse relationship between cerebral cortex and white matter, with higher concentrations in all parts of cerebral cortex compared with cerebral white matter. GFAP, NFL and tau displayed an anteroposterior gradient in the cerebral white matter, with higher concentrations more posteriorly for GFAP and NFL, and the opposite for tau. In the cerebrum, UCHL1 concentration was slightly higher in sections of grey matter (cerebral cortex, caudate nucleus, and hippocampus) than in white matter, but the concentration in the internal capsule, thalamus, brainstem and cervical spinal cord did not differ from the cerebral cortex. Among the studied regions, the cerebellum stands out, with generally low concentrations of all the investigated proteins.
The warm time affected the levels of both MBP and NFL; higher concentrations were associated with increased duration of warm time (Additional file 1: Figs. S3, S4). This could be due to post-mortem cell degradation releasing proteins from cell structures. In a study on post-mortem changes in rat brain, most changes in protein levels occurred after 24 h, and some protein levels increased post-mortem, but not NFL, which in that study decreased [7]. Another study on post-mortem concentration of NFL in human frontal cortex could not confirm any correlation with post-mortem interval [8]. Contrary to our result, MBP concentration decreased with increasing post-mortem interval in a study on vascular white matter changes and dementia [9]. The lack of association between protein concentrations and duration of cold time should be expected, since low temperature retards the post-mortem degradation processes.
To our knowledge, there is no previous study that has systematically quantitated the concentration and distribution of these proteins throughout the CNS. Previous studies on these proteins using ELISA on CNS tissue homogenates are scarce and have focused on a single or a few CNS regions. Petzold et al. reported a GFAP concentration of 1.7 (1.1–5.9) (median, range) µg/mg of total protein in cerebral white matter and 0.8 (0.5–1.2) (median, range) µg/mg of total protein in cerebral cortex from five deceased control patients in a study on multiple sclerosis [10]. In another study, GFAP concentration of 0.8 (0.6–1.1) (median, IQR) µg/mg of total protein in non-lesional white matter in 12 patients with multiple sclerosis was reported [11]. These concentrations of GFAP were normalised to total protein concentration, and were similar to the concentrations in our study. In a study on Alzheimer disease, the concentration of tau in cerebral cortex was 115.8 µg/g tissue in a control subject – again, similar to our findings [12]. The total protein yield was on average 4% (w/w), which is similar with a previous study on drug transporter abundance in post-mortem brain tissue from 30 individuals, using the same protein assay [13].
Extensive open access databases describing the human brain proteome are available, such as the Human Protein Atlas (https://www.proteinatlas.org), or the Allen Institute for Brain Science (https://alleninstitute.org) [5, 14]. The information in these databases relies on gene expression data, immunohistochemistry or in situ hybridization. Our results concerning GFAP, MBP, tau and UCHL1 were consistent with gene expression data from the Human Protein Atlas, less so for NFL. For instance, the highest gene expression of GFAP was reported for mesencephalon, medulla oblongata and spinal cord, which is in line with the actual protein distribution we measured in this study. Another example is cerebral cortex, where MBP was reported to have low and tau high gene expression, which is consistent with our results. NFL on the other hand was in the Human Protein Atlas reported to have a relatively low gene expression in the spinal cord, whereas we found relatively high NFL concentrations in the cervical spinal cord. This inconsistency can be explained by the fact that gene expression data does not reliably reflect the concentration of proteins present in the tissue [15]. Immunohistochemistry on the other hand, gives a good spatial view of the protein distribution but lacks quantitative information. Mass spectrometry allows for (semi) quantification of protein abundance in a tissue, and proteomic profiling of some parts of the human brain have been made [16]. However, the relative abundances of our proteins of interest between the seven investigated brain areas included in that study are not presented, making comparisons difficult to perform. In this study we used ELISA, allowing for absolute protein quantification with high specificity, measuring the protein of interest directly through antibody capture. In comparison with gene expression, mass spectrometry and immunohistochemistry, we believe that our approach better reflects the actual amount present in the tissue, and what eventually will get into the blood when a tissue-damaging process affects a certain part of the CNS.
For many years, analysis of cerebrospinal fluid has been a requirement for obtaining a reliable measurement of these proteins. The development of novel highly sensitive analytical assays has made it possible to reliably quantify the concentrations of these neuroglial proteins in blood samples [17, 18]. The simplicity of a blood test increases the clinical usefulness, and analysis of serum NFL is already approaching clinical routine [19, 20]. Knowledge about the actual distribution of these proteins in the CNS provides a foundation for a correct interpretation of circulating levels in relation to size and location of a CNS damage. Several studies have demonstrated a correlation between infarct size and serum or plasma levels of GFAP, MBP, NFL and tau in acute ischemic stroke [21,22,23,24]. It is reasonable to assume that a stroke affecting mainly white matter would give another pattern (e.g. high serum levels of MBP but not tau) than a stroke affecting mostly grey matter in cortex or basal ganglia (e.g. low serum levels of MBP but high levels of UCHL1 and tau). There are however no studies specifically investigating such patterns and correlation to stroke location. Correlation with lesion volume is seen in other neurological disorders as well; in a recent study of 197 patients with traumatic brain injury, serum measurements of GFAP, NFL, tau and UCHL1 correlated with lesion volume on MRI [3]. In multiple sclerosis, serum NFL correlates with lesion load, lesion volume and gadolinium enhancing lesions on MRI [25, 26]. Another example is serum measurement of GFAP, that is associated with both disease activity and severity in neuromyelitis optica, an inflammatory disorder targeting astrocytes, leading to attacks of longitudinal myelitis and optic neuritis [27]. These examples will most likely be followed by more in the future.
This study has several limitations. The sample size was small, limited to ten donors. Nevertheless, considering the nature of the study this can be considered a fairly large material. We relied upon protein extractions from post-mortem tissue, where factors like post-mortem interval, cause of death and other characteristics possibly could have affected protein concentration. To remedy this, we searched for associations between donor characteristics and protein concentrations and could only find an association between two of the proteins (MBP and NFL) and warm time, which was then adjusted for. Further, GFAP, MBP, NFL and tau have previously been evaluated as post mortem markers of traumatic brain injury, indicating that these proteins remain stable post-mortem [28, 29]. Cause of death was not included in our model due to the limited number of donors, and there is a possibility that causes of death could influence post-mortem concentrations of neuroglial proteins differently. One alternative approach would have been to use biopsy material obtained at neurosurgery. This was not considered feasible, since it would not permit a systematic collection of tissue from standardised CNS regions and also introduce the possibility of confounding by trauma or disease-specific mechanisms.
Although we used ELISA kits from well-known suppliers, these had not been validated for neuronal tissue homogenates, and there is a risk of matrix interference. To minimize interference, samples were diluted between 500 and 1,000,000 times, with a dilution of 10,000 being the most frequent dilution needed for a correct read-off. A high dilution factor reduces the risk of matrix interference.
In conclusion, in this study we presented how five clinically important neuroglial proteins are distributed in the CNS. There was a substantial variation in the concentration of the investigated proteins and between CNS regions. This information is useful when interpreting circulating levels of these proteins in relation to localisation and extent of a CNS-damaging disease.





留言 (0)