
記住我




Acute lung injury is the leading cause of acute respiratory distress syndrome linked to morbidity and death in patients who have suffered trauma (1). Most blunt chest trauma patients are treated conservatively (like about 50% of the patients with a chest tube), as additional approaches to support lung regeneration after direct or indirect trauma remain limited and are primarily centered around immunomodulatory strategies (2–4).
Adult stem and progenitor cells (SPCs) are able to confer immunomodulation or regenerate tissue after injury. Local or circulating SPCs that home to the site of injury support immune-modulation or regeneration by producing soluble factors, such as chemokines, cytokines and growth factors promoting anti-inflammatory and cell-protective effects or stimulating self-regenerative and repair processes or replacing cleared dead cells via differentiation (5). Mesenchymal stroma/stem cells (MSCs) are associated with both immune-modulatory and regenerative potential (6, 7). In lung tissue, it was proposed that MSCs (8) or rare lung progenitor cells are involved in producing type II alveolar cells and thereby improving alveolar repair. Furthermore, Mao et al. (9) suggested that bone marrow-derived endothelial progenitor cells (EPCs), that are able to support re-endotheliation, are recruited to the injured lung to support the restoration of air–blood-barrier functions, indicating a role for local and circulating SPCs in lung regeneration. Exogenously or/and endogenously provided MSCs but also hematopoietic stem and progenitor cells (HSPCs) or EPCs have already been tested for treating bone fractures, liver injuries, burn and wound healing in animal models (10–14) and their use upon trauma has been linked to improved trauma regeneration in mice based on increased vascularization, improvement in the inflammatory cytokine profile and an accelerated healing (15–17).
As of now though, both SPC-based and immunomodulation therapies show only moderate clinical success. Studies to determine at which time point post-trauma SPCs or immune cell modulatory applications show the best therapeutic benefit have not been performed in great detail (18). We hypothesized that SPC-based and immunomodulation therapies in trauma might be further improved by determining the dynamics of mobilization of endogenous SPC after the injury to then target exogenous or endogenous mobilization of SPCs to time points in which the number of endogenously mobilized SPCs is low. Thus, we first characterized the dynamic changes in number in peripheral blood (PB) and the lungs in an established and clinically meaningful murine model of blunt thorax trauma (TXT). We synchronously determined the number of different types of SPCs (three distinct types of MSC populations plus HSPCs plus EPCs) and differentiated hematopoietic cells linked to inflammation up to 48 h post-TXT in PB and the lungs.
To investigate whether therapeutic interventions that increase endogenous SPC or immunomodulatory cells at the site of injury might enhance parameters of TXT outcome, we applied the short-term mobilizer AMD3100 to animals at distinct time points post-trauma and determined inflammatory cytokine levels, plasma markers of organ function, apoptosis, proliferation and the fibroblast/myofibroblast ratio in lung tissue. Our data demonstrate coordinated changes in the number of distinct types of endogenous SPCs and leukocytes post-TXT in blood and other organs, while there is likely only a limited positive role of endogenously mobilized SPCs and immunomodulatory cells on outcome parameters of TXT.
MATERIALS AND METHODS MiceAdult (12 weeks) male C57BL/6 (Janvier Labs, Saint Berthevin, France) were used for all experiments. All mice were housed under specific pathogen-free conditions at the animal housing facility at Ulm University, at a 12:12 night and day cycle with food and water ad libitum. Experiments were performed in compliance with the German Law for Welfare of Laboratory Animals and were approved by the Regierungspräsidium Tübingen (Approval Number 1321).
Blunt chest traumaA single blast injury was applied as previously described (19). Shortly, mice were anesthetized using a 3.5% sevoflurane (AbbVie, Wiesbaden, Germany) and 96.5% oxygen mixture. Buprenorphine (Essex Pharma, Munich, Germany) (Temgesic®) was injected subcutaneously (0.03 mg/kg body weight) before inducing blast injury. Blast injury was applied by opening of a high-speed valve (Hee-D-24, Festo, Esslingen, Germany), delivering compressed air into the upper section of a cylinder. The upper section is separated from the lower section of the cylinder with a polyester film (Mylar A 50mm, Du Pont de Nemur, Bad Homburg, Germany), this polyester film ruptured at a standardized pressure, releasing a reproducible single blast wave toward the nozzle which is centered on the ventral thorax of the animal with a defined distance of 1.5 cm. This set-up induces pulmonary contusion with histological and immunological changes as illustrated in previous studies (19, 20). Sham procedure included analgesia and anesthesia without the blunt chest trauma. Mice were randomly assigned within a cage to be within the sham or TXT group. Mice were monitored for 24 h in which buprenorphine was given twice (0.03 mg/kg body weight). For sample collection mice were sacrificed.
Sample collectionPB was collected by cardiac puncture. To obtain enough progenitor cells, PB of 2 animals was pooled. Mature blood cells were measured with the Hemavet950 (Drew Scientific, France). Lung cells were isolated using the lung dissociation kit (Miltenyi, Bergisch Gladbach, Germany) according to the protocol provided by Miltenyi. In brief, lungs were flushed through the right heart ventricle with 3 mL to 5 mL of PBS. After removal of external tissues, single lung lobes were infiltrated with the enzyme solution and lung cells were isolated using the gentle MACS Octo Dissociator with heater. After isolation, red cell lysis was performed. To obtain total BM, femora and tibiae were flushed and cell count was taken by a Hemavet950. Mononucleated cells from the bone marrow (LDBM) and peripheral blood (LDPB) were isolated by centrifugation on Histopaque-1083 (Sigma-Aldrich, St. Louis, United States). In PB and BM cell numbers were measured by Hemavet950.
Flow cytometry Mature hematopoietic cell stainingMarkers and gating to identify mature hematopoietic cells in the lung is illustrated in Figure S1a, https://links.lww.com/SHK/B440. Shortly, FC-Block anti-mouse CD16/CD32 Clone 93 (eBioscience, San Diego, CA) was added to 106 lung cells, followed by the antibody mixture: anti-CD45, anti-CD11b, anti-CD11c, anti-CD64, anti-CD24, anti-IE/IA, anti-Ly6g, and anti-Ly6c (Table S1, https://links.lww.com/SHK/B440). Before analyzing (BD LSRII, Becton Dickinson, Franklin Lakes, NJ), propidium iodide (PI) (1:1000 Sigma-Aldrich) for live/dead staining was applied.
HSPC and EPC stainingA biotin-labeled lineage-cocktail was added to LDPB cells, to 2 × 106 LDBM cells and to 3 × 106 lung cells (Table S1, https://links.lww.com/SHK/B440). For the staining, anti-CD45, streptavidin, anti-CD34, anti-Sca-1, anti-c-Kit, and anti-Flk-1 were applied for 1 h at 4°C in the dark (Table S1, https://links.lww.com/SHK/B440). After incubation, cells were washed and before measuring by flow cytometry (BD LSRII, Becton Dickinson, Franklin Lakes, NJ) PI was added for live/dead separation. The BM staining served as gating control for PB and lung (Fig. S1b/c, https://links.lww.com/SHK/B440). An additional size gate for the Lin-Sca-1+c-Kit+ (LSK) cells in lung tissue was applied to reduce number of false positive cells (see Fig. S1b, https://links.lww.com/SHK/B440).
MSC stainingBiotin-labeled anti-CD45, anti-CD31, and anti-lineage-cocktail was added to LDPB cells and to 2 × 106 LDBM cells (Table S1, https://links.lww.com/SHK/B440). For the staining, streptavidin, anti-CD44, anti-Sca-1, anti-CD29, anti-CD51, anti-CD90, and anti-CD105 were applied for 1 h at 4°C in the dark (Table S1, https://links.lww.com/SHK/B440). After incubation, cells were washed and analyzed by flow cytometry (BD LSRII). BM was used as a gating control. The MSC 2 marker panel is based on MSC detection kit mouse (Novus) and the MSC 3 marker panels are based on MSC marker kit from Abcam. Gating strategy is illustrated in Figure S1d, https://links.lww.com/SHK/B440.
Annexin V and PI stainingLung cells (2 × 106) were incubated with Viability dye eFluor780 (1:1000) (eBioscience, San Diego, CA) for 30 min. Then the samples were washed and stained in 1× binding buffer plus Annexin V-V450 antibody. Before analyzing by flow cytometry (LSRII, Becton Dickinson, Franklin Lakes, NJ) PI was added to a final concentration of 50 μg/mL, to allow for quantification of cell cycle distribution.
Fibroblast/myofibroblast stainingLung cells (2–3 × 106) were incubated with non-fibroblast lineage cocktail (Table S1, https://links.lww.com/SHK/B440). Cells were washed and FC-Block was added. Followed by an AB-incubation with anti-Sca-1, anti-CD49e, and Streptavidin for 1 h at 4°C, samples were measured by flow cytometry (LSRII, Becton Dickinson, Franklin Lakes, NJ). Gating was applied as previously described (21).
All flow cytometric data were analyzed using BD FACSDIVA software 8.1, obtained cell numbers were used for the calculation of the absolute cell number. For the calculation of the fold change, cage mates underwent sham procedure and were sacrificed at the same time points, absolute cell number in TXT was divided through absolute cell number in sham to reduce batch to batch variations.
AMD3100 treatmentMice were treated with 5 mg/kg AMD3100 (Sigma-Aldrich, St. Louis, United States) which was freshly dissolved in PBS or PBS only (PAN-Biotech, Aidenbach, Germany). Solutions were injected intraperitoneally (i.p.) directly after or 6 h post-TXT induction.
ELISAFrozen lung tissue was homogenized and sonicated. Levels of interleukin-6 (IL-6), IL-10, keratinocyte chemoattractant (KC), monocyte chemotactic protein-1 (MCP-1), and tumor necrosis factor-α (TNF-α) (all R&D Systems, Minneapolis, United States) were measured by serial ELISA. Blood plasma was collected and stored at −80°C. Levels of heart-type fatty acid binding protein (H-FABP) (Life Diagnostics, West Chester, United States), Syndecan-1 (Biozol, Eching, Germany), and Clara cell secretory protein (CC16) (Abbexa, Cambridge, Great Britain) were determined by ELISA.
Data analysis and statisticsData were checked for normal distribution with the Shapiro–Wilk normality test. Cell numbers and mobilization data in PB and lung were compared to a fold change of one, which indicates no differences between sham and TXT. For normal distributed data the one sample t test and for non-normally distributed the Wilcoxon signed-rank test was applied. For multiple comparison data normally distributed was analyzed using one-way analysis of variance (ANOVA), followed by Tukey's multiple comparison test, data non-normally distributed was analyzed by the Kruskal–Wallis test, followed by Dunn's multiple comparison test. The significance level was determined at 5% (∗P ≤ 0.05, ∗∗P ≤ 0.01, ∗∗∗P ≤ 0.001, ∗∗∗∗P ≤ 0.0001). An a priori power analysis to determine the number of biological repeats (n) was performed that guided the experimental set-up. Data are displayed as mean ± standard deviation (SD). All statistical analyses were performed with Graph Pad prism 7 software (GraphPad Software, San Diego, CA).
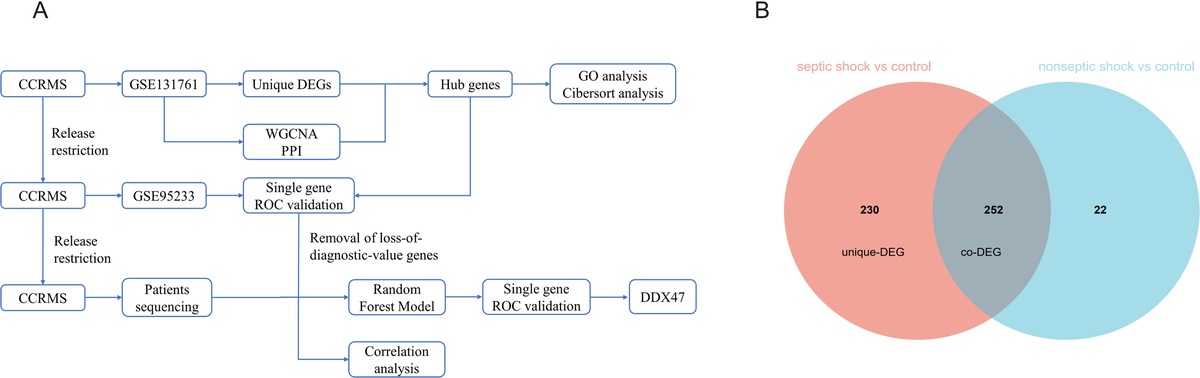
RESULTS The frequencies of SPCs in PB and lung change upon TXTWe first determined the change in the frequency of HSPCs and EPCs in PB and lung between 0.2 and 48 h post-TXT by flow cytometry (gating strategy Fig. S1b, c, https://links.lww.com/SHK/B440). We observed a slight nadir for the frequency of HSCPs in PB at 6 and 12 h post-TXT compared to controls, while 48 h post-TXT the frequency of late hematopoietic progenitors and HSCs in PB were elevated (1.4- and 2-fold) (Fig. 1A). In the lung, the frequency of hematopoietic progenitors and HSCs remained similar up to 24 h post-TXT while it was also increased for early hematopoietic progenitors and HSCs (2- and 3.2-fold) at 48 h after TXT (Fig. 1B). The frequency of EPCs (Flk-1+, CD34+, c-Kit+, tested for endothelial potential, Fig. S1c, https://links.lww.com/SHK/B440) was also modestly altered in PB upon TXT (1.9-fold at 12 h and 1.7-fold at 24 h post-TXT) (Fig. 1A) but interestingly not in the lung (Fig. 1B). Changes in the frequency of HSPCs as well as EPCs among tissues did not correlate and did thus not change in a real synchronized fashion (Table S2, https://links.lww.com/SHK/B440). Protocols for the identification of murine MSCs are, in contrast to protocols for human MSCs, quite heterogeneous with respect markers. We therefore applied three published but distinct marker panels for the identification of MSCs (Fig. S1d, https://links.lww.com/SHK/B440, MSC 1: Lin−, CD31−, CD45−, CD51+(22) and Sca-1+; MSC2: Lin−, CD31−, CD45−, CD29+, CD105+, and Sca-1; MSC 3: Lin−, CD31−, CD45−, CD29+, CD90+, CD44+, and Sca-1+). Furthermore, the frequency of MSCs could only be determined in PB, as the MSCs marker panels are also expressed on non-MSCs of the lung. The change in frequency of MSC 1 did correlate with the change in frequency of MSC 3 (Spearman's r = 0.52, P = 0.0009). MSC types 1 and 3 showed an increase in their frequency in PB at 2 h post-TXT (3- and 2.2-fold), and all three MSC populations at 24 h post-TXT (Fig. 1C).
 Fig. 1:
Fig. 1: Fold change (TXT/sham) in the tissue content of different types of SPCs over time after TXT. (A–B) HSPC and EPC number changes in (A) PB; n = 5–7, (B) lung injured tissue; n = 5–7; (C) MSC number changes in PB; n = 3–7; (D, E) Inflammatory cell number changes in (D) PB; n = 5–7 and (E) lung injured tissue; n = 4–7. Data are presented as mean ± SD; one sample t test for normal distributed time points and Wilcoxon signed-rank test for non-normal distributed time points was preformed to the fold change of one, ∗P≤0.05, ∗∗P≤0.01, ∗∗∗P≤0.001. Outliers were identified and removed by ROUT, with Q = 1%.
HSPCs, EPCs as well as MSCs frequency in PB are at their lowest 6 and 12 h post-TXT, while they are increased compared to the normal level at 2 h as well as 24 to 48 h post-TXT. A nadir of HSPCs in PB was also observed in polytrauma (PT) patients, although at 24 h post-PT. The overall dynamics of the frequency of SPCs in mice upon TXT is therefore somewhat similar to the inverse bell-shaped pattern of the frequency of HSCPs in PB of polytrauma patients that was recently described by us (23), but with an accelerated change in mice.
Innate inflammatory cells are recruited early upon TXT to PB and lung tissueWe next quantified the change in the frequency of lymphocytes, monocytes and neutrophils (cells linked to the inflammatory response upon TXT) in PB and lung between 0.2 and 48 h post-TXT (Fig. 1D, E). The frequency of these cells in PB and lung was elevated 2 h post-TXT (lymphocytes 1.8-fold, monocytes 3.3-fold, and neutrophils 4.1-fold in PB, and 2.1-, 4.6-, and 13.1-fold in lung tissue). Of note, 6 h post-injury leukocytosis was already resolved, indicating an initially fast resolution of the inflammatory response. Neutrophils showed another increase in frequency (2.3-fold) 24 h post-TXT in PB and (2.9-fold) 48 h post-TXT also in lung tissue. Similar to SPCs, the number of leukocytes was at its lowest in PB and the lung at 6 h post-TXT.
Pharmacological inhibition of Cxcr4 induces a coordinated mobilization of HSPCs and leukocytes in TXT animalsAMD3100 blocks the Cxcl12/Cxcr4 axis, leading to a mobilization of various types of cells including SPCs into PB and other tissues. Whether AMD3100 also results in a coordinated increase in the frequency of SPCs and leukocytes in TXT animals in PB as well as lung tissue to positively influence TXT outcome parameters has not been systematically investigated. To this end, AMD3100 was injected right after TXT induction and/or 6 h post-TXT (experimental setup, Fig. 2A). Mobilization with AMD3100 peaks at 1 to 2 h post-application. As there is a strong increase of leukocytes in PB at 2 h post-TXT even without further cell mobilization, providing AMD3100 right after TXT will test the influence of even further enhanced numbers of leukocytes as well as SPCs in blood very early after TXT on outcome. In contrast, as we observed the nadir of the number of SPCs and leukocytes in blood around 6 h post-TXT, AMD3100 will then likely increase the number of SPCs in the circulation to at least regular levels to test whether alleviation of this nadir might change outcomes.
 Fig. 2:
Fig. 2: Experimental overview and early fold change in cell numbers after TXT and administration of AMD3100 or PBS. (A) Experimental overview of the endpoint analysis after TXT induction and AMD3100 or PBS treatment. At 2 and 6 h two experimental groups (AMD3100 or PBS treatment at 0 h after TXT induction), at 24 and 48 h four experimental groups (PBS treatment at 0 and 6 h, AMD3100 treatment at 0 and 6 h, AMD3100 treatment at 0 h/PBS treatment at 6 h and PBS treatment at 0 h/AMD3100 treatment at 6 h. (B, C) Early fold change (2 h) after the application of TXT and application of AMD3100 or PBS. Fold change is calculated by dividing the absolute cell number in TXT + AMD3100 treated animals by the absolute cell number in TXT+PBS treated animals. (B) PB and (C) lungs; n = 3–4; data are presented as mean ± SD; one sample t test for normal distributed cell populations and Wilcoxon signed-rank test for non-normal distributed cell populations was preformed to the fold change of one, ∗P≤0.05, ∗∗P≤0.01.
AMD3100 did indeed result in a strong increase in the number of leukocytes (lymphocytes, monocytes, and neutrophils, fold change of 2.5, 4.0, and 3.0), HSCs (9.7-fold), early progenitors (9.1-fold), and late hematopoietic progenitors (4.9-fold) and EPCs (5.8-fold) after 2 h in PB. However, none of the MSCs populations showed a significant increase in frequency 2 h post-AMD3100 compared to non-mobilized TXT animals (Fig. 2B). In lung tissue, the frequency of neutrophils (3.8-fold) and monocytes (5.1-fold) was increased in response to AMD3100, but not that of lymphocytes. In contrast, the frequency of HSPCs was only marginally increased in comparison to control TXT animals, while the frequency of EPC was even slightly decreased (0.5-fold) (Fig. 2C). Taken together, AMD3100 increases the frequency of HSPCs and neutrophils and monocytes in PB and neutrophils and monocytes in lung tissue compared to TXT control animals.
Mobilization of SPCs and leukocytes by AMD3100 does not affect parameters of TXT outcomeTo test the hypothesis that endogenously mobilized SPCs might exert a positive influence, molecular and tissue markers of TXT outcome were analyzed. We determined the concentration of inflammatory cytokines in the lung, plasma markers of organ function and the level of apoptosis, proliferation and the ratio of myofibroblasts over fibroblasts in the lung.
TXT induces a localized inflammation with an upregulation in the pro-inflammatory cytokines (19, 20, 24, 25). Although AMD3100 caused an increase in neutrophils and monocytes in the lung (Fig. 2C), the concentration of the pro-inflammatory cytokines IL-6, MCP-1, TNF-α, KC and the anti-inflammatory cytokine IL-10 in lung tissue at 6 and 24 h post-TXT remained similar in TXT animals treated with AMD3100 relative to control TXT animals treated only with PBS (Fig. 3A, B).
 Fig. 3:
Fig. 3: Cytokine levels and fibroblast recruitment in lung after TXT. (A) Inflammatory cytokine levels in lung tissue at 6 h after TXT and AMD3100 treatment; n = 5–7; mean ± SD; Mann–Whitney test. (B) Cytokine and chemokine measurement in homogenized lung tissue 24 h after TXT application and AMD3100 treatment(s) (− = PBS was applied; + = AMD3100 was applied); n = 5–7; mean ± SD; Kruskal–Wallis test (Dunn's multiple comparison test).
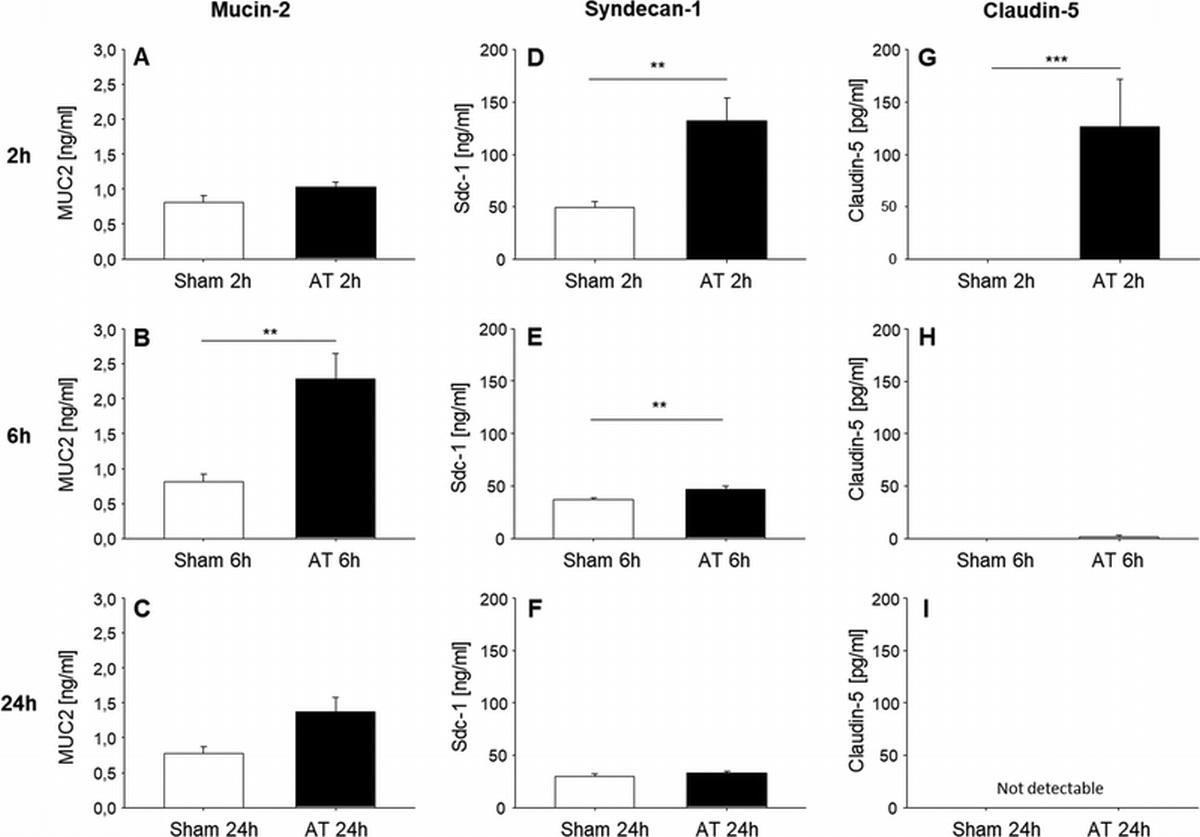
Additionally, we measured the level of plasma markers of TXT target organ function such as CC16, Syndecan-1, and H-FABP in response to AMD3100 induced mobilization. CC16, released from the non-ciliated bronchiolar Clara cells, is an accepted blood biomarker for the detection of direct and indirect lung injury (26), whereas Syndecan-1 can be used as a plasma biomarker of endothelial damage and was for example shown to be associated with burn severity and subsequent level of vascular permeability in lung of rodents (27). H-FABP is an established plasma marker for cardiac cell damage. We included this marker due to findings that TXT induces cardiac inflammation and structural alterations in cardiac tissue in rodents (28). TXT animals treated with AMD3100 did not show significant changes in the level of these markers at either the 6 h or 24 h timepoint (Fig. 4A/B), further supporting the notion that AMD3100 induced mobilization of SPCs has likely only a limited positive influence on the initial TXT regeneration process.
 Fig. 4:
Fig. 4: Circulating damage markers in plasma and myofibroblast ratio, proliferation and apoptosis in lung after TXT. (A) Circulating damage markers in plasma 6 h after TXT and AMD3100 treatment; n = 5–7; mean ± SD; unpaired t test. (B) Circulating damage markers in plasma 24 h after TXT application and AMD3100 treatment(s) (− = PBS was applied; + = AMD3100 was applied); n = 5–7; mean ± SD; one-way ANOVA (Tukey's multiple comparisons test). (C) Myofibroblast/fibroblast ratio in lung 48 h after TXT. Left panels: Representative example of gating for the identification of fibroblasts (Fibro.) and myofibroblasts (Myofibro.) Right panel: Ratio between the percentage of fibroblasts and myofibroblasts (− = PBS treatment; + = AMD3100 treatment); n = 8; mean ± SD; Kruskal–Wallis test (Dunn's multiple comparison test). (D) Apoptotic cells were identified by Annexin V staining 48 h after TXT induction and AMD3100/PBS treatment(s); n = 5–6; mean ± SD; one-way ANOVA (Tukey's multiple comparisons test). (E) Fold change % of proliferating cells 48 h after TXT and different treatment(s) (− = PBS treatment, + = AMD3100 treatment) in lung tissues; n = 5–6; mean ± SD; one-way ANOVA (Tukey's multiple comparisons test).
Upon TXT, the number of myofibroblasts in the lung increases. Prolonged persistence of myofibroblast after trauma is linked to tissue fibroses and scaring (29). We thus determined the change in fibroblasts/myofibroblasts ratio (Fig. 4C, left, (21)) upon TXT and in response to AMD3100 induced SPC and leukocyte mobilization in TXT. The frequency of myofibroblasts and the ratio of fibroblasts/myofibroblasts remained similar in TXT animals treated with AMD3100 compared to TXT controls (Fig. 4C, right). The regeneration progress of TXT is also characterized by both apoptosis and proliferation. The frequency of apoptotic cells as well as the level of cell proliferation was also not altered in AMD3100 treated TXT animals (Fig. 4D, E).
In summary, AMD3100 induced mobilization of SPCs and neutrophils and monocytes did neither influence the inflammatory response nor regeneration parameters in the lung of TXT animals.
DISCUSSIONStem cell-based therapies are promising approaches in regenerative medicine. Here, we hypothesized that endogenous SPCs are mobilized to PB and the site of injury at distinct time points post-trauma to likely affect trauma outcome, and that providing elevated levels of endogenously SPCs at distinct time point post-trauma via mobilization might improve detrimental consequences of trauma. Similar to our analyses in human polytraumatized (PT) patients (26), we observed a trend toward an inverse bell-shaped pattern for the frequency of HSPCs and EPCs in PB after TXT, with a modest increase of their number 2 h and a nadir at 6 h post-trauma, followed by an increase at the later time points. Whether a more severe murine trauma model system that is more similar to human PT than the applied TXT model might show SPCs dynamics in mice even more similar to human polytrauma thus warrants further investigations.
Interestingly, changes in the frequency of SPC within lung were negligible upon TXT, with the exception of an increase in the number of HSCs 48 h post-trauma. As HSPCs can support regeneration by providing effector cells and are able to release paracrine factors that stimulate tissue repair and modulate the immune response and influence vascularization (12, 17, 30), it is a possibility that HSCs in the lung 48 h post-trauma might indeed contribute to lung regeneration.
The short-time mobilizer AMD3100 effectively mobilized, in a coordinated fashion, HSPCs, EPCs, neutrophils, and monocytes also in TXT animals, but surprisingly not MSCs. Considering the important role of the Cxcl12/Cxcr4 axis in the regulation of MSC migration, we expected an increase of their number in PB upon mobilization in TXT. Toupadakis et al. (10) for example induced a higher number of MSCs in PB after AMD3100 treatment in a bone fracture model. Liu et al. (31) in contrast could only increase the number of MSCs in PB by pre-treating animals with the hypoxia-mimicking agent cobalt chloride but not by AMD3100 alone.
In contrast to PB, we detected no increase in the frequency of HSPCs in the lung of TXT animals after AMD3100 mobilization. It is a possibility that the Cxcl12/Cxcr4 axis is still partly blocked by AMD3100 at 2 h post-injection (plasma half live of 2–3 h (32)), which might actually mute migration of these cells (as this might be require a functional Cxcl12/CXCR4 axis) into the lung. We further found even a reduction of the number of EPCs in the lung upon AMD3100 application. Yin et al. (33) showed that while AMD3100 stimulates mobilization it also reduces EPC functionality in part by inducing apoptosis, which might contribute to the reduction of EPCs in the lung upon AMD3100 administration.
With respect to TXT regeneration, mobilization of SPCs and leukocytes remained neutral for all the established outcome parameters analyzed, which is in contrast to some published reports indicating beneficial effects of for example exogenously provided SPCs on trauma outcome (34). In aggregation, our data support the absence of a strong positive effect of AMD3100 induced mobilization on outcome parameters of TXT, while they also support that additional research will be required to unequivocally determine the magnitude of the contribution of endogenously mobilized SPC or leukocytes to TXT outcome.
Limitations of the studyOur sample size used in the experiments here is too small to allow for a proper validation of the significance of small differences in outcome parameters. It thus remains a possibility that additional experiments based on much larger samples size might identify a positive, but then likely smaller effect of AMD3100 induced mobilization on TXT outcome parameters. However, the biological relevance of small changes in for example levels of cytokines with respect to change in regeneration after TXT remain in general more difficult to interpret.
AcknowledgmentsWe thank the Tierforschungszentrum University Ulm for their help with mouse work and housing of the mice. Additionally, we thank Anne Rittlinger and Anke Schultze for excellent technical support. Furthermore, we thank all the members of the SFB1149 for the stimulating discussions and helpful comments.
REFERENCES 1. Huber-Lang M, Lambris JD, Ward PA. Innate immune responses to trauma. Nat Immunol 19 (4):327–341, 2018. 2. Chrysou K, Halat G, Hoksch B, Schmid RA, Kocher GJ. Lessons from a large trauma center: impact of blunt chest trauma in polytrauma patients—still a relevant problem? Scand J Trauma Resusc Emerg Med 25 (1):42, 2017. 3. Julier Z, Park AJ, Briquez PS, Martino MM. Promoting tissue regeneration by modulating the immune system. Acta Biomater 53:13–28, 2017. 4. Lord JM, Midwinter MJ, Chen YF, Belli A, Brohi K, Kovacs EJ, Koenderman L, Kubes P, Lilford RJ. The systemic immune response to trauma: an overview of pathophysiology and treatment. Lancet 384 (9952):1455–1465, 2014. 5. Pati S, Pilia M, Grimsley JM, Karanikas AT, Oyeniyi B, Holcomb JB, Cap AP, Rasmussen TE. Cellular therapies in trauma and critical care medicine: forging new frontiers. Shock 44 (6):505–523, 2015. 6. Ren G, Zhang L, Zhao X, Xu G, Zhang Y, Roberts AI, Zhao RC, Shi Y. Mesenchymal stem cell-mediated immunosuppression occurs via concerted action of chemokines and nitric oxide. Cell Stem Cell 2 (2):141–150, 2008. 7. Selvasandran K, Makhoul G, Jaiswal PK, Jurakhan R, Li L, Ridwan K, Cecere R. A tumor necrosis factor-α and hypoxia-induced secretome therapy for myocardial repair. Ann Thorac Surg 105 (3):715–723, 2018. 8. Hayes M, Curley GF, Masterson C, Devaney J, O’Toole D, Laffey JG. Mesenchymal stromal cells are more effective than the MSC secretome in diminishing injury and enhancing recovery following ventilator-induced lung injury. Intensive Care Med Exp 3 (1):29, 2015. 9. Mao S-Z, Ye X, Liu G, Song D, Liu SF. Resident endothelial cells and endothelial progenitor cells restore endothelial barrier function after inflammatory lung injury. Arterioscler Thromb Vasc Biol 35 (7):1635–1644, 2015. 10. Toupadakis CA, Granick JL, Sagy M, Wong A, Ghassemi E, Chung DJ, Borjesson DL, Yellowley CE. Mobilization of endogenous stem cell populations enhances fracture healing in a murine femoral fracture model. Cytotherapy 15 (9):1136–1147, 2013. 11. Seebach C, Henrich D, Wilhelm K, Barker JH, Marzi I. Endothelial progenitor cells improve directly and indirectly early vascularization of mesenchymal stem cell-driven bone regeneration in a critical bone defect in rats. Cell Transplant 21 (8):1667–1677, 2012. 12. Si Y, Tsou CL, Croft K, Charo IF. CCR2 mediates hematopoietic stem and progenitor cell trafficking to sites of inflammation in mice. J Clin Invest 120 (4):1192–1203, 2010. 13. Oh EJ, Lee HW, Kalimuthu S, Kim TJ, Kim HM, Baek SH, Zhu L, Oh JM, Son SH, Chung HY, et al. In vivo migration of mesenchymal stem cells to burn injury sites and their therapeutic effects in a living mouse model. J Control Release 279:79–88, 2018. 14. Li M, Luan F, Zhao Y, Hao H, Liu J, Dong L, Fu X, Han W. Mesenchymal stem cell-conditioned medium accelerates wound healing with fewer scars. Int Wound J 14 (1):64–73, 2017. 15. Wang XX, Allen RJ, Tutela JP, Sailon A, Allori AC, Davidson EH, Paek GK, Saadeh PB, McCarthy JG, Warren SM. Progenitor cell mobilization enhances bone healing by means of improved neovascularization and osteogenesis. Plast Reconstr Surg 128 (2):395–405, 2011. 16. Putra A, Antari AD, Kustiyah AR, Intan YSN, Sadyah NAC, Wirawan N, Astarina S, Zubir N, Munir D. Mesenchymal stem cells accelerate liver regeneration in acute liver failure animal model. Biomed Res Ther 5 (11):2802–2810, 2018. 17. Allakhverdi Z, Delespesse G. Hematopoietic progenitor cells are innate Th2 cytokine-producing cells. Allergy 67 (1):4–9, 2012. 18. Pati S, Rasmussen TE. Cellular therapies in trauma and critical care medicine: looking towards the future. PLoS Med 14 (7):e1002343, 2017. 19. Knöferl MW, Liener UC, Seitz DH, Perl M, Brückner UB, Kinzl L, Gebhard F. Cardiopulmonary, histological, and inflammatory alterations after lung contusion in a novel mouse model of blunt chest trauma. Shock 19 (6):519–525, 2003. 20. Hafner S, Wagner K, Wepler M, Matallo J, Gröger M, McCook O, Scheuerle A, Huber-Lang M, Frick M, Weber S, et al. Physiological and immune-biological characterization of a long-term murine model of blunt chest trauma. Shock 43 (2):140–147, 2015. 21. Akamatsu T, Arai Y, Kosugi I, Kawasaki H, Meguro S, Sakao M, Shibata K, Suda T, Chida K, Iwashita T. Direct isolation of myofibroblasts and fibroblasts from bleomycin-injured lungs reveals their functional similarities and differences. Fibrogenesis Tissue Repair 6 (1):15, 2013. 22. Guidi N, Sacma M, Ständker L, Soller K, Marka G, Eiwen K, Weiss JM, Kirchhoff F, Weil T, Cancelas JA, et al. Osteopontin attenuates aging-associated phenotypes of hematopoietic stem cells. EMBO J 36 (7):840–853, 2017. 23. Vogel M, Christow H, Manz I, Denkinger M, Amoah A, Schütz D, Brown A, Möhrle B, Schaffer A, Kalbitz M, et al. Distinct dynamics of stem and progenitor cells in blood of polytraumatized patients. Shock 51 (4):430–438, 2019. 24. Perl M, Gebhard F, Braumüller S, Tauchmann B, Brückner UB, Kinzl L, Knöferl MW. The pulmonary and hepatic immune microenvironment and its contribution to the early systemic inflammation following blunt chest trauma. Crit Care Med 34 (4):1152–1159, 2006. 25. Niesler U, Palmer A, Fröba JS, Braumüller ST, Zhou S, Gebhard F, Knöferl MW, Seitz DH. Role of alveolar macrophages in the regulation of local and systemic inflammation after lung contusion. J Trauma Acute Care Surg 76 (2):386–393, 2014. 26. Broeckaert F, Bernard A. Clara cell secretory protein (CC16): characteristics and perspectives as lung peripheral biomarker. Clin Exp Allergy 30 (4):469–475, 2000. 27. Cruz MV, Carney BC, Luker JN, Monger KW, Vazquez JS, Moffatt LT, Johnson LS, Shupp JW. Plasma ameliorates endothelial dysfunction in burn injury. J Surg Res 233:459–466, 2019. 28. Kalbitz M, Amann EM, Bosch B, Palmer A, Schultze A, Pressmar J, Weber B, Wepler M, Gebhard F, Schrezenmeier H, et al. Experimental blunt chest trauma-induced myocardial inflammation and alteration of gap-junction protein connexin 43. PLoS ONE 12 (11):e0187270, 2017. 29. Glasser SW, Hagood JS, Wong S, Taype CA, Madala SK, Hardie WD. Mechanisms of lung fibrosis resolution. Am J Pathol 186 (5):1066–1077, 2016. 30. Rafii S, Butler JM, Ding BS. Angiocrine functions of organ-specific endothelial cells. Nature 529 (7586):316–325, 2016. 31. Liu L, Yu Q, Fu S, Wang B, Hu K, Wang L, Hu Y, Xu Y, Yu X, Huang H. CXCR4 antagonist AMD3100 promotes mesenchymal stem cell mobilization in rats preconditioned with the hypoxia-mimicking agent cobalt chloride. Stem Cells Dev 27 (7):466–478, 2018. 32. Jujo K, Hamada H, Iwakura A, Thorne T, Sekiguchi H, Clarke T, Ito A, Misener S, Tanaka T, Klyachko E, et al. CXCR4 blockade augments bone marrow progenitor cell recruitment to the neovasculature and reduces mortality after myocardial infarction. Proc Natl Acad Sci USA 107 (24):11008–11013, 2010. 33. Yin Y, Huang L, Zhao X, Fang Y, Yu S, Zhao J, Cui B. AMD3100 mobilizes endothelial progenitor cells in mice, but inhibits its biological functions by blocking an autocrine/paracrine regulatory loop of stromal cell derived factor-1 in vitro. J Cardiovasc Pharmacol 50 (1):61–67, 2007. 34. Amann EM, Rojewski MT, Rodi S, Fürst D, Fiedler J, Palmer A, Braumüller S, Huber-Lang M, Schrezenmeier H, Brenner RE. Systemic recovery and therapeutic effects of transplanted allogenic and xenogenic mesenchymal stromal cells in a rat blunt chest trauma model. Cytotherapy 20 (2):218–231, 2018.
留言 (0)