
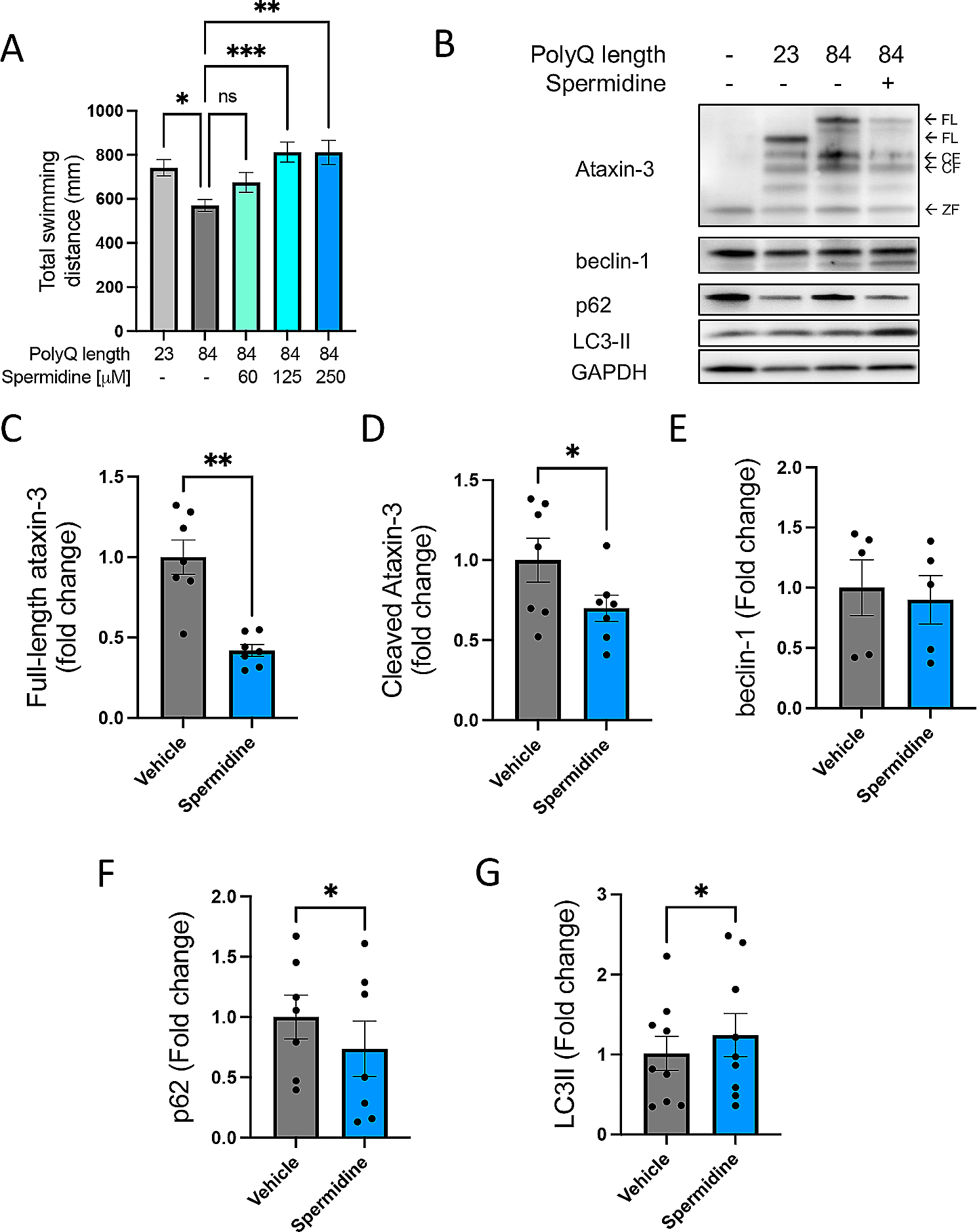
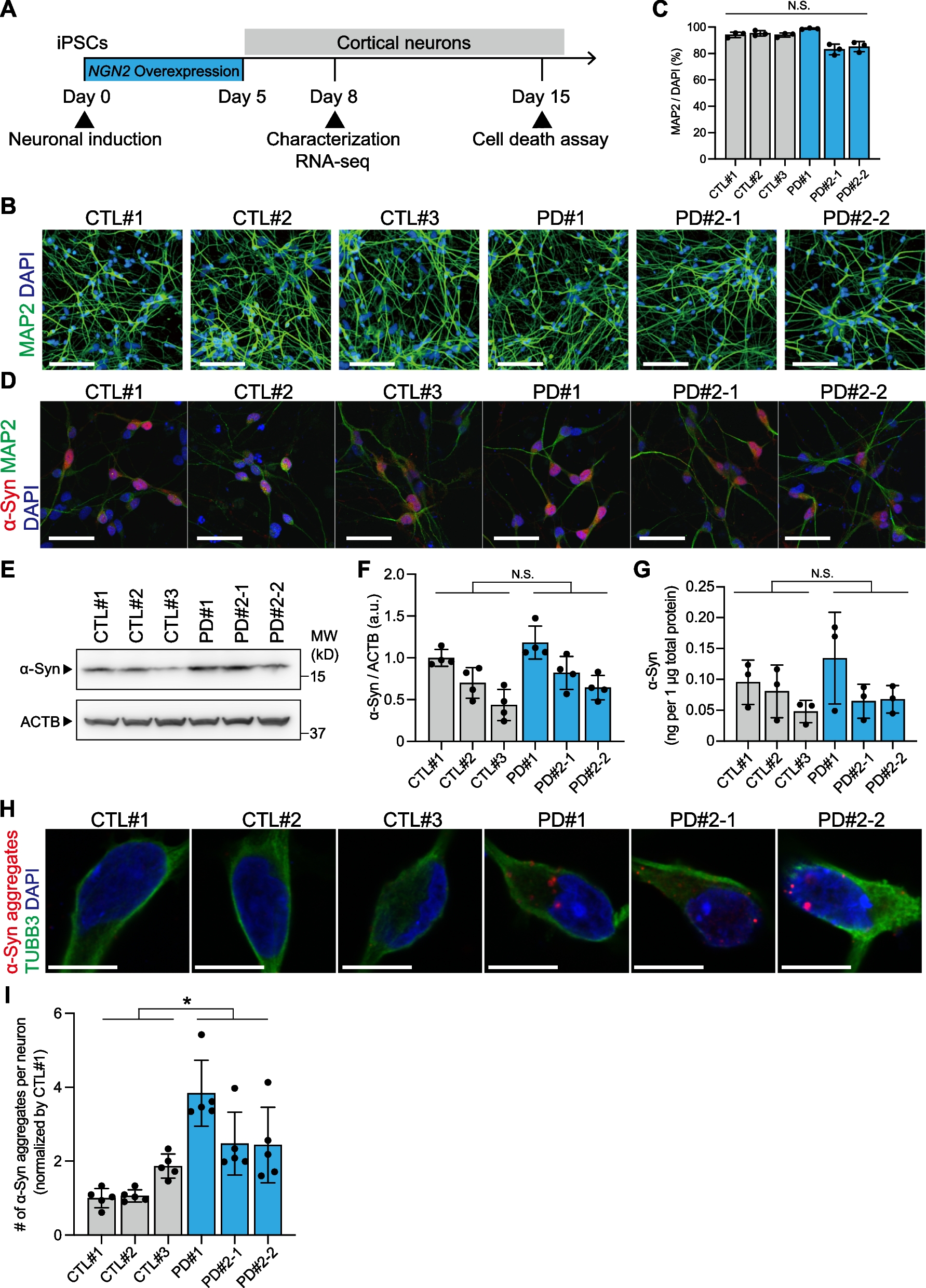
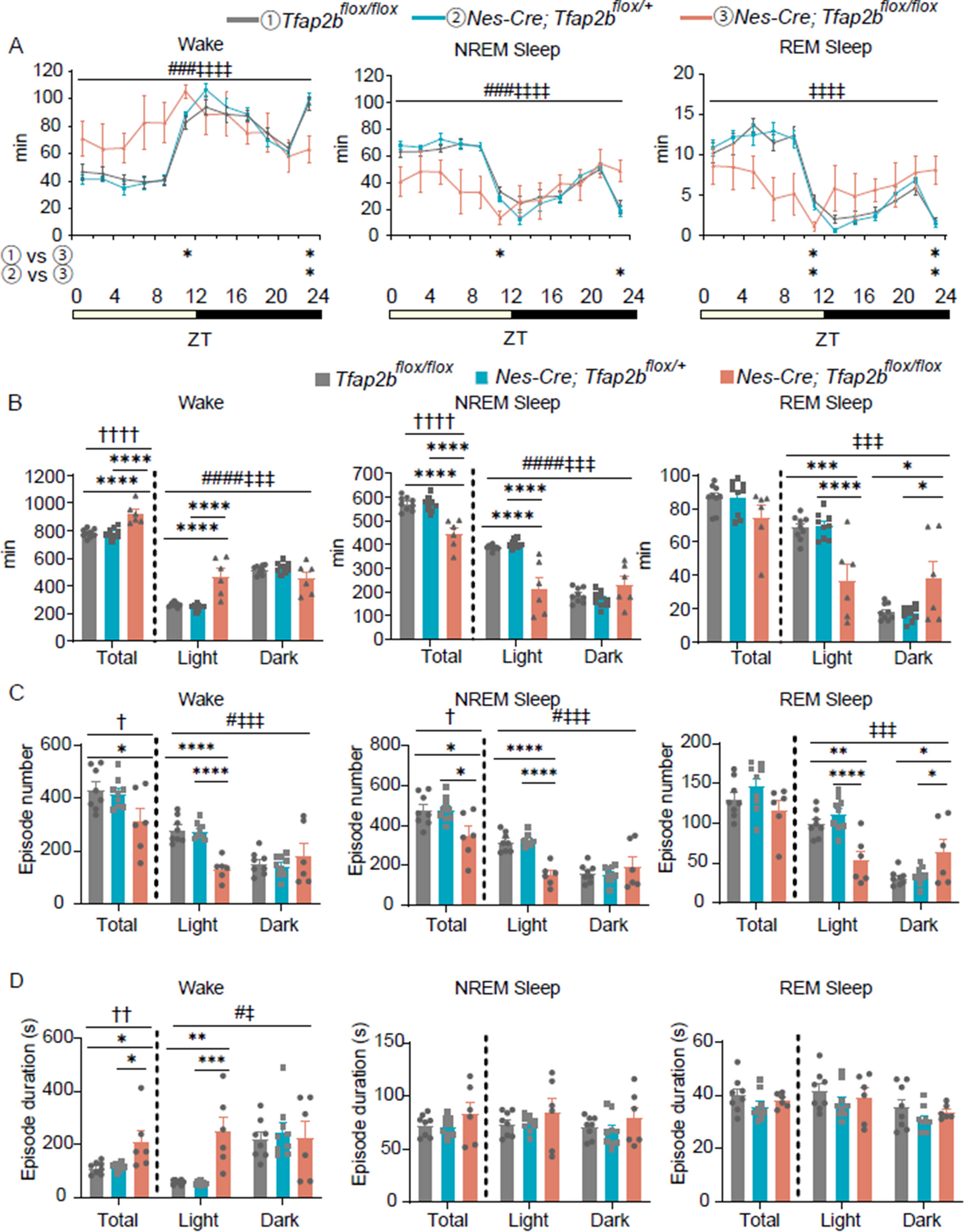
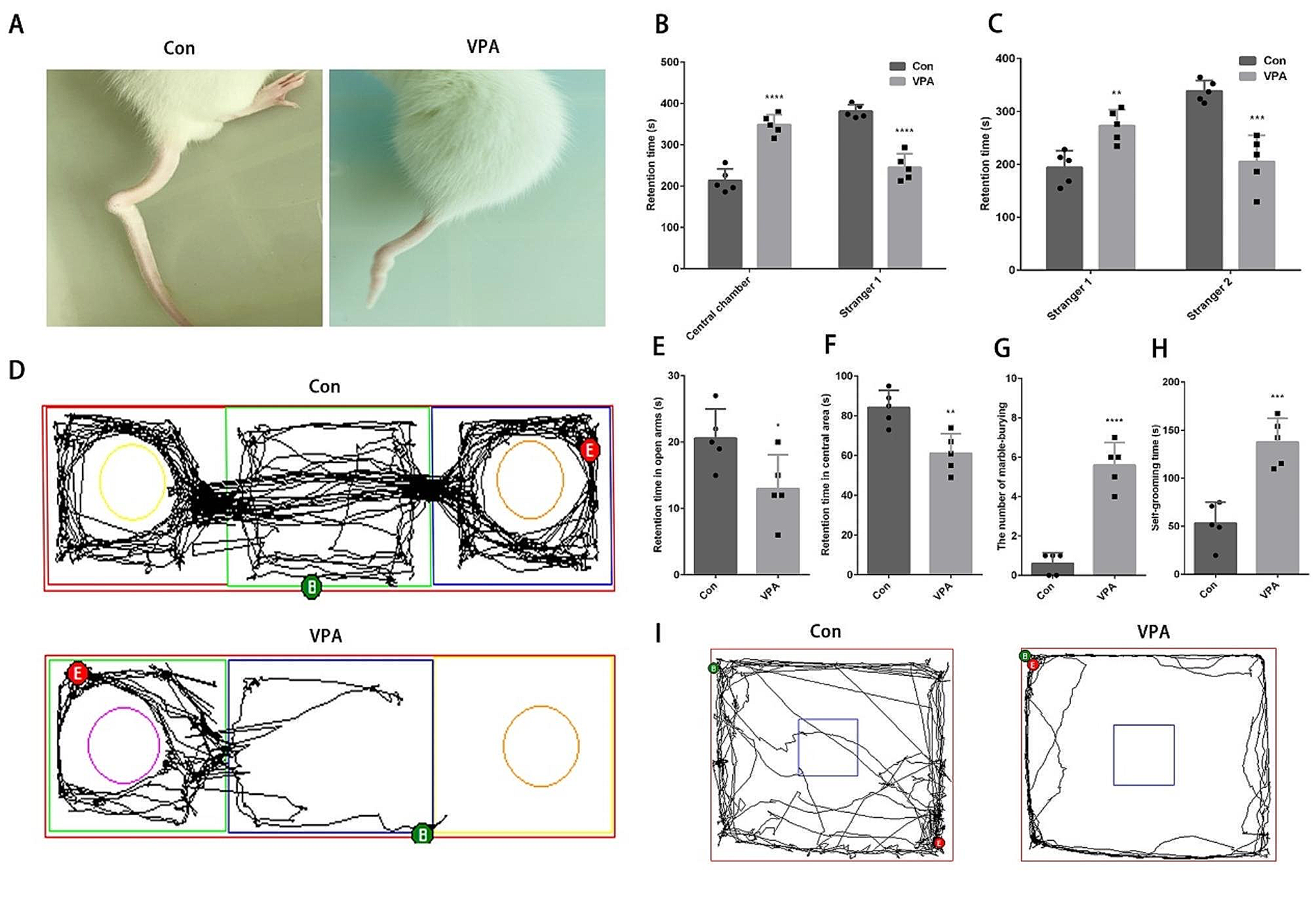
記住我



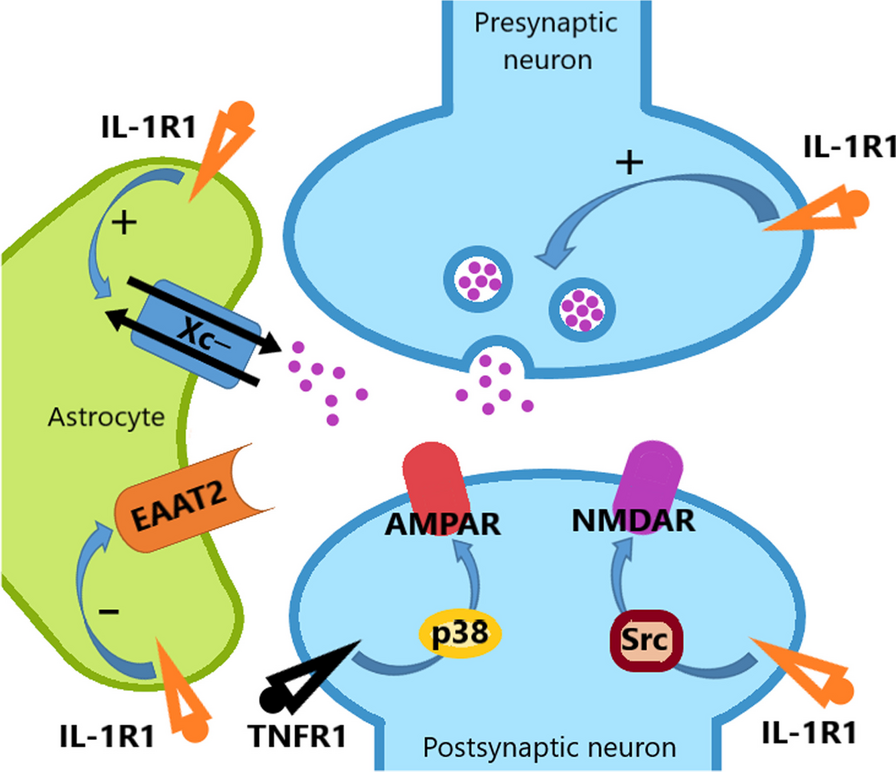

First, we verified that deleting Rptor specifically in PV cells affects Raptor expression in hippocampal CA1 PV interneurons. We quantified the number of EYFP-expressing hippocampal CA1 PV INs that are immunopositive for Raptor in PV-Raptor-WT, PV-Raptor-Het and PV-Raptor-Homo mice. We found that the number of PV INs expressing EYFP and immunopositive for Raptor are significantly reduced in PV-Raptor-Homo mice relative to PV-Raptor-WT mice (PV-Raptor-WT n = 8 sections, PV-Raptor-Het n = 6 sections, PV-Raptor-Homo n = 7 sections, for 3 mice in each group; One Way ANOVA, F(2, 18) = 4 p < 0.0001, Tukey’s multiple comparisons test, PV-Raptor-WT vs PV-Raptor-Het, p = 0.5; PV-Raptor-WT vs PV-Raptor-Homo, p < 0.0001; PV-Raptor-Het vs PV-Raptor-Homo, p < 0.0001; Fig. 1A, B). PV-Raptor-Het mice failed to show such a decrease in number of PV INs expressing EYFP and immunopositive for Raptor. Thus, Raptor expression is impaired in hippocampal PV INs of PV-Raptor-Homo mice.
Fig. 1
Homozygous conditional knock-out of Rptor in PV interneurons causes a deficit in mTORC1 activity. A Left, representative images of EYFP-positive PV interneurons (white arrows) in PV-Raptor-WT, PV-Raptor-Het and PV-Raptor-Homo mice injected with AAV2/9-DIO-EYFP in dorsal CA1 hippocampus. Right, representative images of Raptor-positive (red) EYFP-expressing PV interneurons (white arrows, co-labeling) in PV-Raptor-WT, PV-Raptor-Het and PV-Raptor-Homo mice. Scale bar 100 µm. B Summary graph showing reduced percentage of EYFP-positive cells that are also Raptor-positive in PV-Raptor-Homo mice relative to PV-Raptor-Het and PV-Raptor-WT mice (n = 7 sections from 3 PV-Raptor-WT mice, 6 sections from 3 PV-Raptor-Het mice, and 7 sections from 3 PV-Raptor-Homo mice, from 3 independent experiments). C Representative images showing EYFP-positive cells (green), S6S240/244 phosphorylation (red) and co-labeling (merged) in CA1 hippocampus of PV-Raptor-WT, PV-Raptor-Het and PV-Raptor-Homo mice. D Summary graph showing reduced p-S6 immunofluorescence in CA1 PV interneurons of PV-Raptor-Homo mice relative to PV-Raptor-Het and PV-Raptor-WT mice (n = 5 mice/group from 5 independent experiments, scale bar 50 µm). **** p < 0.0001, * p < 0.05, ns not significant
Next, we examined the effects of conditional knock-out of Rptor on mTORC1 activity by assessing phosphorylation of ribosomal protein S6S240/244 (p-S6), a downstream effector of mTORC1, in CA1 PV INs using immunofluorescence. The level of p-S6 was reduced in EYFP-expressing CA1 PV INs of PV-Raptor-Homo mice relative to PV-Raptor-WT or PV-Raptor-Het mice (n = 5 mice in each group; one-way ANOVA, F (2, 12) = 8, p = 0.006; Tukey’s multiple comparisons test, PV-Raptor-WT vs PV-Raptor-Het, p = 0.99; PV-Raptor-WT vs PV-Raptor-Homo, p = 0.01; PV-Raptor-Het vs PV-Raptor-Homo, p = 0.01; Fig. 1C, D). The level of p-S6 was unaffected in CA1 PV INs of PV-Raptor-Het mice relative to PV-Raptor-WT mice. These results confirm that mTORC1 signaling, as assessed by p-S6, is impaired in hippocampal PV INs of PV-Raptor-Homo mice.
Conditional Rptor knock-out in PV interneurons impairs repetitive firingActivation of mTORC1 is generally linked to stimulation of protein synthesis [27, 38, 39]. However, mTORC1 activation also represses the synthesis of specific mRNAs, such as the Kv1.1 channel, a voltage-gated potassium channel that regulates neuronal excitability [40, 41]. Therefore, we determined if conditional Rptor knock-out in PV INs affected their membrane and firing properties. Whole-cell patch-clamp recordings were obtained from EYFP-expressing PV INs located in or near the CA1 stratum pyramidale in acute slices from control and PV conditional Rptor knock-out mice (PV-Raptor-WT n = 9 cells in 4 mice, PV-Raptor-Het n = 10 cells in 3 mice, and PV-Raptor-Homo n = 11 cells in 3 mice). We found that PV INs from control and conditional knock-out mice had similar resting membrane potential (One Way ANOVA, F (2, 27) = 0.65 p = 0.52; Fig. 2A) and input resistance (One Way ANOVA, F(2, 27) = 1.93 p = 0.16; Fig. 2B), suggesting intact basic membrane properties.
Fig. 2
Conditional knock-out of Rptor in PV interneurons impairs firing properties. A and B Summary bar graphs showing intact: resting membrane potential (A) and input resistance (B) of CA1 PV interneurons from PV-Raptor-Het (green) and PV-Raptor-Homo (red) relative to PV-Raptor-WT mice (black). C Representative voltage responses of PV interneurons in response to a somatic depolarization (260 pA), illustrating the decrease in evoked firing of PV cells from PV-Raptor-Het and PV-Raptor-Homo mice relative to PV-Raptor-WT mice. D Summary plot of frequency-current relationship for all cells showing reduced evoked firing of PV interneurons from PV-Raptor-Homo (n = 11 cells, 3 mice) and PV-Raptor-Het (n = 10 cells, 3 mice) mice relative to PV-Raptor-WT mice (n = 9 cells, 4 mice). E–I Summary bar graphs showing intact: action potential amplitude (E), action potential latency (F), action potential threshold (G), action potential half-width (H) and fast after-hyperpolarization amplitude (I) of CA1 PV interneurons from PV-Raptor-Het (green) and PV-Raptor-Homo (red) relative to PV-Raptor-WT mice (black). * p < 0.05, **** p < 0.0001, ns not significant
Then, we assessed PV interneurons repetitive firing properties in response to somatic depolarizations. PV INs from control and mutant mice responded to incremental somatic depolarization with increasing number of action potentials (Fig. 2C, D). However, PV INs from PV-Raptor-Het and PV-Raptor-Homo mutant mice fired less action potentials compared to those from PV-Raptor-WT mice (Two Way ANOVA, F (2, 27) = 7.73 p = 0.002; Tukey’s multiple comparisons tests, PV-Raptor-WT vs. PV-Raptor-Het p < 0.0001, PV-Raptor-WT vs PV-Raptor-Homo p < 0.0001, PV-Raptor-Het vs. PV-Raptor-Homo p = 0.012; Fig. 2C, D). The impairment in firing was greater in PV INs from PV-Raptor-Het mice compared to those from PV-Raptor-Homo mice. These results suggest that conditional hetero- and homozygous knock-out of Rptor in PV interneurons have impaired firing output.
Next, we examined whether the firing impairment of PV INs could be explained by changes in action potential properties. We found that PV INs from PV-Raptor-Het and PV-Raptor-Homo mice display similar action potential amplitude (One Way ANOVA, F(2, 27) = 0.44 p = 0.65, Fig. 2E), latency to first action potential (One Way ANOVA, F(2, 27) = 2.60 p = 0.09 Fig. 2F), action potential threshold (One Way ANOVA, F(2, 27) = 02.89 p = 0.07, Fig. 2G), action potential half-width (One Way ANOVA, F(2, 27) = 0.54 p = 0.58, Fig. 2H) and fast afterhyperpolarization amplitude (One Way ANOVA, F(2, 27) = 0.64 p = 0.53, Fig. 2I). Together, these data indicate that conditional hetero- and homozygous knock-out of Rptor in PV interneurons impairs their repetitive firing properties without affecting their resting membrane potential, input resistance and action potential properties.
Conditional Rptor knock-out in PV interneurons impairs long-term potentiation of intrinsic excitabilityHippocampal GABAergic interneurons are highly dynamic and display several forms of long-term plasticity of synapses and intrinsic excitability [14, 15, 19,20,21, 42]. CA1 parvalbumin-expressing basket cells show long-term potentiation of intrinsic excitability (LTPIE) via mGluR5 activation and down-regulation of Kv1.1 channels, which is prevented by rapamycin, an inhibitor of mTORC1 [19]. Given that Raptor is obligatory for mTORC1 function, we tested if conditional knock-out of Rptor in PV INs could affect LTPIE. We obtained whole-cell patch-clamp recording from EYFP-positive PV interneurons located in or near CA1 stratum pyramidale. After establishing their fast-spiking phenotype in current-clamp mode via the injection of depolarizing currents, we adjusted (i) the intracellular depolarizing current to evoke approximately 5 action potentials, and (ii) the extracellular electrode simulation in stratum radiatum to elicit an EPSP of approximately 2 mV in amplitude [19]. After obtaining a stable baseline (5 min) of depolarization-evoked firing, we applied a high frequency stimulation (HFS) to the Schaffer collaterals pathway that consisted of 10 pulses at 100 Hz, repeated 10 times at the frequency of 3 Hz, and recorded PV interneuron spiking induced by the same somatic depolarization for up to 30 min [19].
As previously reported in rat [19], we found that in PV INs from PV-Raptor-WT mice (n = 10 cells in 6 mice), HFS of Schaffer collaterals resulted in long-lasting potentiation of PV IN evoked firing (159.8% ± 15.8% of baseline at 10–15 min and 197.3% ± 3.3% of baseline at 25–30 min post-HFS, paired t-tests, p = 0.013 and p = 0.011 respectively, Fig. 3A, B). In the absence of HFS of Schaffer collateral pathway, we observed no change in evoked firing of PV INs (n = 5 cells from 5 mice, 105.18% ± 14% of baseline at 10–15 min and 118.65% ± 15% of baseline at 25–30 min post-HFS, paired t-tests, p = 0.88 and p = 0.21 respectively, Fig. 3A, B). LTPIE was associated with a reduction in the latency of the first action potential (79.89 ± 5.49% of baseline at 10–15 min post-HFS and 68.17 ± 6.67% of baseline at 25–30 min post-HFS, Wilcoxon tests, p = 0.004 and 0.004 respectively, Fig. 3C) and a hyperpolarization of the first action potential threshold (102.8 ± 0.54% of baseline at 10–15 min post-HFS and 106.54 ± 1.16% of baseline at 25–30 min post-HFS, paired t-tests, p = 0.0007 and 0.0003 respectively, Fig. 3D). In the absence of HFS of Schaffer collateral pathway, no change was observed in the first action potential latency (97.13 ± 14.73% of baseline at 10–15 min post-HFS and 91.8 ± 15.23% of baseline at 25–30 min post-HFS, paired t-tests, p = 0.83 and 0.55 respectively, Fig. 3C), and only a depolarization of the first action potential threshold was seen at 10–15 min post-HFS (95.04 ± 0.7% of baseline at 10–15 min post-HFS and 98.2 ± 2.76% of baseline at 25–30 min post-HFS, paired t-tests, p = 0.0018 and 0.55 respectively, Fig. 3D). The reduction in the first spike latency and threshold in the HFS group but not in the No HFS group is consistent with a modulation of Kv1.1 during LTPIE in PV interneurons, as previously reported [19]. Overall, these data confirm that HFS of Schaffer collateral pathway causes LTPIE in PV interneurons, which is dependent on tetanization, not due to unspecific effects of recording conditions and is associated with a modulation of Kv1.1 channels.
Fig. 3
Conditional knock-out of Rptor in PV interneurons impairs LTPIE. A Representative traces (top) and time plot for all cells (bottom) of depolarization evoked firing showing long-lasting increase of firing in the group receiving HFS (filled circles, HFS) but not in the non-tetanized control group (open circles, No HFS) in CA1 PV interneuron from PV-Raptor-WT mice. B-D Summary plots of spikes number (B), latency to first spike (C) and first spike threshold (D) measured at -5 to 0 min baseline versus 10–15 min (left), or 25–30 min (right) post-HFS in cells of the tetanized group (HFS, top) and control group (No HFS, bottom). Individual data points before and after are joined by lines; means ± sem are indicated to the side for each group. E–H Similar data representation showing absence of long-lasting potentiation of intrinsic excitability at 10–15 min and 25–30 min after HFS in PV interneurons from PV-Raptor-Het mice. I–L Similar data representation showing a block of long-lasting potentiation of intrinsic excitability at 10–15 min, but not at 25–30 min, after HFS in PV interneurons from PV-Raptor-Homo mice. M and N Summary plots of spike increases relative to baseline for all cells measured at 10–15 min (M) and 25–30 min (N) after HFS in PV interneurons, showing block of LTPIE at 10–15 and 25–30 min after HFS in PV-Raptor-Het mice, and at 10–15 min post HFS in PV-Raptor-Homo mice, relative to PV-Raptor-WT mice. *** p < 0.001, ** p < 0.01, * p < 0.05, ns not significant
Next, we assessed whether the conditional knock-out of Rptor in PV interneurons affected LTPIE since it was reported to be sensitive to the mTORC1 inhibitor rapamycin [19]. In PV-Raptor-Het mice, we found that HFS of Schaffer collaterals failed to induce long-term potentiation of evoked firing in PV mice (n = 13 cells from 5 mice, 91.1% ± 14% of baseline at 10–15 min and 105.5% ± 23.1% of baseline at 25–30 min post-HFS, paired t-tests, p = 0.67 and p = 0.8 respectively, Fig. 3E, F). HFS also failed to alter consistently the first action potential latency (99.66 ± 3.26% of baseline at 10–15 min post-HFS and 98.98 ± 7.33% of baseline at 25–30 min post-HFS, paired t-tests, p = 0.09 and 0.43 respectively, Fig. 3G) and threshold (101.53 ± 1% of baseline at 10–15 min post-HFS and 103.22 ± 1.24% of baseline at 25–30 min post-HFS, paired t-tests, p = 0.15 and 0.02 respectively, Fig. 3H). As in PV-Raptor-WT mice, the absence of HFS stimulation in PV-Raptor-Het mice did not affect PV interneuron evoked firing (n = 5 cells from 3 mice, 85.11% ± 9.6% of baseline at 10–15 min and 88.9% ± 20% of baseline at 25–30 min post-HFS, paired t-tests, p = 0.16 and p = 0.37 respectively, Fig. 3E, F), latency of first action potential (91.86 ± 6.73% of baseline at 10–15 min post-HFS and 91.1 ± 8.35% of baseline at 25–30 min post-HFS, paired t-tests, p = 0.23 and 0.31 respectively, Fig. 3G) or threshold of first action potential (102.66 ± 1.57% of baseline at 10–15 min post-HFS and 99.27 ± 2.27% of baseline at 25–30 min post-HFS, paired t-tests, p = 0.20 and 0.76 respectively, Fig. 3H). These data suggest that conditional heterozygous deletion of Rptor in PV interneurons is sufficient to prevent LTPIE.
In PV-Raptor-Homo mice, HFS failed to induce a potentiation of evoked firing at 10–15 min but did elicit an increase in firing at 25–30 min post-HFS (n = 14 cells from 5 mice, 115.53% ± 10% of baseline at 10–15 min and 144.9% ± 18.7% of baseline at 25–30 min post-HFS, paired t-tests, p = 0.23 and p = 0.04 respectively, Fig. 3I, J). Similarly, HFS failed to alter latency of the first action potential at 10–15 min but not at 25–30 min post-HFS (97.67 ± 9.64% of baseline at 10–15 min post-HFS, paired t-test, p = 0.4, and 80.95 ± 6.95% of baseline at 25–30 min post-HFS, Shapiro–Wilk test, p = 0.008, Fig. 3K). HFS reduced threshold of the first action potential at 10–15 min but not at 25–30 min post-HFS (102.62 ± 1.28% of baseline at 10–15 min post-HFS and 102.86 ± 1.5% of baseline at 25–30 min post-HFS, paired t-tests, p = 0.04 and p = 0.07 respectively, Fig. 3L). These results suggest that LTPIE is impaired by homozygous deletion of Rptor, but that an mTORC1-independent late component of LTPIE remains, as previously reported in experiments using the mTORC1 inhibitor rapamycin [19]. In the absence of HFS, PV interneuron from PV-Raptor-Homo mice did not show change over the same time period in evoked firing (n = 6 cells from 3 mice, 100.3% ± 11% of baseline at 10–15 min and 99.9% ± 20% of baseline at 25–30 min post-HFS, paired t-tests, p = 0.56 and p = 0.72 respectively, Fig. 3I, J), latency to first action potential (90.06 ± 9.35% of baseline at 10–15 min post-HFS, paired t-test, p = 0.64 and 104.66 ± 16.60% of baseline at 25–30 min post-HFS, Shapiro–Wilk test, p = 0.68 respectively, Fig. 3K) and threshold of first action potential (100.15 ± 1.58% of baseline at 10–15 min post-HFS and 100.05 ± 1.6% of baseline at 25–30 min post-HFS, paired t-tests, p = 0.95 and p = 0.98 respectively, Fig. 3L), confirming stable evoked firing over the recording period in these mice also.
Overall, these results show that (i) HFS of Schaffer collaterals induces LTPIE in PV interneurons of PV-Raptor-WT mice, (ii) LTPIE is blocked at both 10–15 min and 25–30 min post-HFS in PV-Raptor-Het mice, and (iii) LTPIE is deficient at 10–15 min but not 25–30 min post-HFS in PV-Raptor-Homo mice (Fig. 3M, 10–15 min post-HFS: One way ANOVA, F (2, 34) = 6 p = 0.003, Tukey’s multiple comparisons test, PV-Raptor-WT vs PV-Raptor-Het, p = 0.004, PV-Raptor-WT vs PV-Raptor-Homo, p = 0.036, PV-Raptor-Het vs PV-Raptor-Homo, p = 0.53; Fig. 3N, 25–30 min post-HFS: One way ANOVA, F(2, 34) = 3 p = 0.04, Tukey’s multiple comparisons test, PV-Raptor-WT vs PV-Raptor-Het, p = 0.03, PV-Raptor-WT vs PV-Raptor-Homo, p = 0.27, PV-Raptor-Het vs. PV-Raptor-Homo, p = 0.48). These findings indicate that the hetero- and homozygous conditional knock-out of Rptor in PV INs impairs LTPIE, consistent with previous report that this plasticity is sensitive to the mTORC1 inhibitor rapamycin [19].
Mice with conditional Rptor knock-out in PV interneurons show normal contextual fear memory and fear discriminationLTP of PV interneuron excitatory synapses and coherence of PV interneuron firing with CA1 network oscillations are required for contextual fear memory consolidation [35, 36]. Since we found a deficit in LTPIE in PV interneurons with conditional Rptor knock-out, next we examined fear memory consolidation and discrimination in these mice. During contextual fear conditioning, PV-Raptor-Het and PV-Raptor-Homo mice showed similar freezing responses to foot shocks relative to PV-Raptor-WT mice (n = 21 PV-Raptor-WT mice, 13 PV-Raptor-Het mice, and 15 PV-Raptor-Homo mice; Two way ANOVA F (2, 46) = 0.358, p = 0.7, Fig. 4A, B), indicating normal anxiety and sensorimotor gating in the mutant mice.
Fig. 4
Conditional knock-out of Rptor in PV interneurons does not affect contextual fear memory or context discrimination. A Diagram of the contextual fear memory and context discrimination protocol. B Percentage of time freezing after each foot shock during the training session for PV-Raptor-WT (n = 21), PV-Raptor-Het (n = 13) and PV-Raptor-Homo (n = 15) mice (Baseline: before the first foot shock), indicating similar anxiety level and sensorimotor gating in the three groups. C Percentage of time freezing during the long-term memory tests at 24 h in the PV-Raptor-WT, PV-Raptor-Het and PV-Raptor-Homo mice (same mice as in B), indicating similar long-term contextual memory in the three groups. D–F Percentage of time freezing during the contextual discrimination test, relative to the training context (D), in the new neutral context (E) and discrimination ratio (F; amount of freezing in [training context]/[training context + neutral context]) for PV-Raptor-WT (n = 12), PV-Raptor-Het (n = 7) and PV-Raptor-Homo (n = 6) mice, indicating similar context discrimination in the three mice groups. **** p < 0.0001, *** p < 0.001, * p < 0.05, ns not significant
During the long-term memory test in the training context (24 h after conditioning), PV-Raptor-Het and PV-Raptor-Homo mice showed similar freezing responses relative to PV-Raptor-WT mice (One way ANOVA, F (2, 46) = 0.5923 p = 0.5572, Fig. 4A–C), indicating intact long-term contextual memory in the mutant mice. During the context discrimination test in a new neutral context, the three mice groups showed reduced freezing responses relative to the training context, indicating significant contextual discrimination (paired t-tests; n = 12 PV-Raptor-WT mice, p < 0.0001; n = 7 PV-Raptor-Het mice, p = 0.0026; n = 6 PV-Raptor-Homo mice, p = 0.026; Fig. 4D). In the neutral context, PV-Raptor-Het and PV-Raptor-Homo mice showed similar freezing responses relative to PV-Raptor-WT mice (One way ANOVA, F (2, 22) = 0.4621 p = 0.6359, Fig. 4E). Similarly, discrimination ratios to assess context discrimination normalized to the freezing level in the training context were similar in control and mutant mice (One way ANOVA, F (2, 22) = 2.83 p = 0.08, Fig. 4F). These results suggest that long-term contextual fear memory and context discrimination are intact in mice with conditional Rptor knock-out in PV INs, and, thus, mTORC1 regulation of firing and long-term potentiation of intrinsic excitability of PV INs may not be necessary for long-term contextual fear memory and context discrimination.
留言 (0)