
記住我



An ever-increasing number of researchers emphasise that OPG may not only be a marker of an unfavourable prognosis in CVDs, but may also play an important pathogenetic role in their development. It has been confirmed that OPG is present in vascular ECs and in VSMCs under basic conditions, and that its production increases significantly in these cells under the influence of various stimulating factors, such as cytokines or hormones [4, 6, 17, 32]. In human aorta cells, it was confirmed that the production of OPG is 20–30 times higher in VSMCs than in ECs [6]. TNF-alpha and interleukin-1beta (IL-1B) are strong stimulators of OPG production in ECs and VSMCs. These cytokines can increase OPG expression 5–40-fold [4, 6, 32]. In ECs, OPG occurs in secretory granules, called Weibel-Palade bodies, where OPG is associated with the von Willebrand factor [4, 6].
OPG can exert its biological effect on the CV system in three ways. The first way is that it binds through a specific domain to its ligand RANKL, thereby blocking it from binding to its receptor, RANK. The second mode of action of OPG is to act directly on cells via a heparin-binding domain that has the ability to bind to heparan sulphate proteoglycans present on the surface of cells, triggering cell-surface signalling. Such OPG action has been confirmed in the area of vascular wall cells, bones and cells of the immune system [3, 6, 17, 33]. A third way in which OPG works is when it attaches to its other ligand, TRAIL. This blocks the possibility of TRAIL binding to its receptors, which weakens or abolishes the effects of TRAIL [6].
The RANK-RANKL-OPG signalling axis and the direct action of OPG on cellsMore and more is becoming known about the elements of the RANK-RANKL-OPG axis and the role of the transcription factor and nuclear factor kappa-B (NF-kB) within the ECs and their role in the pathogenesis of inflammation within the vascular wall and atherogenesis [4].
Human RANK is a protein consisting of 616 amino acids. Each of these molecules consists of the C-terminal cytoplasmic domain of 383 amino acids, a signal peptide of 28 amino acids, a transmembrane domain of 21 amino acids and an N-terminal extracellular domain of 184 amino acids [4]. RANK is a transmembrane receptor whose cytoplasmic domain is capable of binding to TNF receptor–associated factors (TRAFs 2,5,6). The binding of RANK to its ligand RANKL, which exists both as a transmembrane protein and in a soluble form (RANKLs) circulating in the blood, results in the activation of a signalling cascade activating the transcription factor NF-kB. RANKL binds to RANK as a homotrimer. The first effect of binding RANK to RANKL is the attachment of TRAFs: 2,5,6 to specific sites within the cytoplasmic domain of RANK. This triggers several signalling pathways activated by RANK/TRAF-mediated protein kinase signalling, such as inhibitory NF-kB kinase (IKK)/NF-kB and activator protein-1 (AP-1). NF-kB and AP-1 acting at the level of the cell nucleus increase the expression of OPG and adhesion molecules such as intercellular adhesion molecule 1 (ICAM-1) and vascular adhesion molecule 1 (VCAM-1), among others. It has been confirmed that OPG also activates NF-kB in ECs and thus induces the expression of ICAM-1 and VCAM-1 at the level of the cell nucleus, which leads to increased adhesion of leucocytes to the endothelial surface in the early stages of endothelial dysfunction [4, 5, 17] (Fig. 2). OPG has been shown to greatly enhance the adherence of leucocytes to the surface of endothelial cells through its heparin-binding domain. OPG binds via this domain to the sulphate proteoglycans present on the surface of an endothelial cell. This connection activates signalling pathways inside the cell, leading, inter alia, to an escalation of the adhesion of leucocytes to the EC surface [17]. In ECs, the sensors and effectors of shear stress also regulate the expression of genes regulated by NF-kB, such as the genes for ICAM-1 and VCAM-1 [4, 34]. Increased shear stress activates NF-kB, which activates gene expression for said particles at the level of the cell nucleus. However, so far, there is no confirmation that intensified shear stress increases the expression of OPG in vascular ECs. It has been confirmed, nevertheless, that shear stress increases the expression of OPG in osteocytes and reduces the influence of interleukin 17A (IL-17A) on the expression of TNF-alpha and RANKL, thus weakening the differentiation of osteoclasts [4].
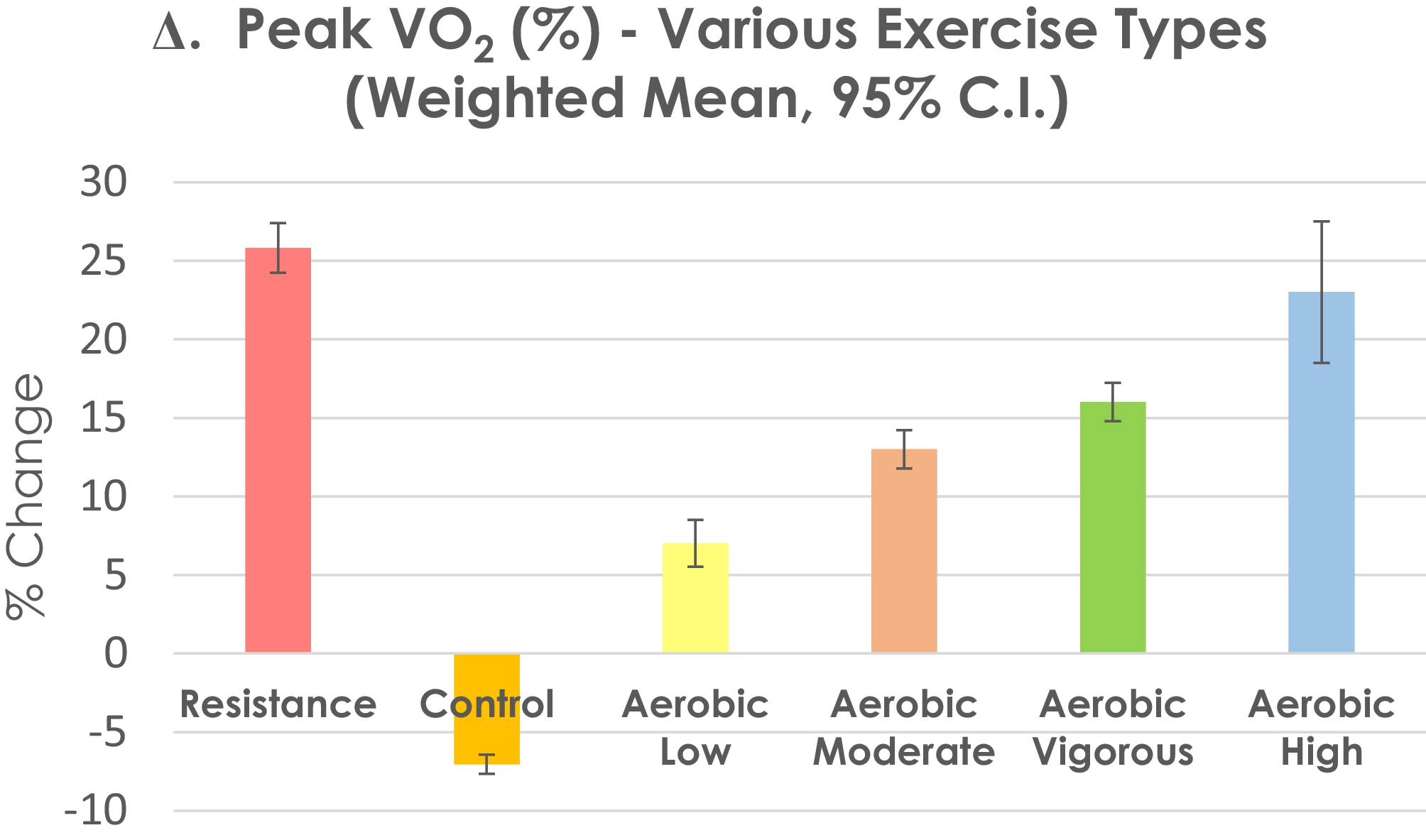
Fig. 2
Mechanisms of action of OPG and TRAIL in endothelial cells. OPG exerts its biological effect on the endothelial cells in three ways. Firstly, it binds through a specific domain to its ligand RANKL, preventing it from binding to its receptor, RANK. Secondly, OPG acts directly on ECs via a heparin-binding domain that has the ability to bind to heparan sulphate proteoglycans present on the surface of cells, triggering cell-surface signalling. Thirdly, it attaches to its other ligand, TRAIL, preventing TRAIL from binding to its receptors, thereby weakening or abolishing its effects. The binding of RANK to its ligand RANKL results in the activation of a signalling cascade activating NF-kB and AP-1. Acting at the level of the cell nucleus, these molecules increase the expression of OPG and ICAM-1 and VCAM-1. OPG also activates NF-kB in ECs inducing the expression of ICAM-1 and VCAM-1 at the level of the cell nucleus, which leads to increased adhesion of leucocytes to the endothelial surface in the early stages of endothelial dysfunction. OPG greatly enhances the adherence of leucocytes to the surface of ECs through its heparin-binding domain. An important connection also exists between OPG and RAS. Activation of the AT1 receptors for Ang II causes, inter alia, an increase in the expression of VEGFS. VEGF-A and VEGF-B increase inflammation and remodelling in blood vessels by activating pro-inflammatory mechanisms and pathological angiogenesis, which are strengthened by OPG. Mutual stimulating interactions between OPG and RAS have also been confirmed. In ECs Ang II increases OPG expression and OPG increases the expression of the AT1 receptor for Ang II. TRAIL significantly increases the activity of eNOS and increases NO production in ECs. The augmented phosphorylation of eNOS and the increase in NO production in ECs under the influence of TRAIL occur through the activation of the PI3 kinase/Akt pathway. Additionally, TRAIL induces PGE2 production in ECs, mainly by increasing COX-1 activity. This effect of TRAIL is accomplished by increasing NO release. NO inhibits the activity of NF-kB and decreases the expression of ICAM-1, VCAM-1 and E-selectin in ECs and prevents increased adherence of leucocytes to the endothelial surface. Explanation of abbreviations in the main text
In addition, low-density lipoproteins (LDLs) accumulating under the endothelial layer, and especially their oxidised forms (OxLDLs), which are involved in the induction of the atherosclerotic process at an early stage, have the ability to stimulate the expression of ICAM-1 and VCAM-1. This contributes to an escalation in the adherence of leucocytes to the endothelial surface. The degree of LDL oxidation is highly dependent on the reactive oxygen species (ROS) generated by the monocytes. The opposite effect is displayed by nitric oxide (NO), which is generated in ECs with the involvement of endothelial nitric oxide synthase (eNOS) and reduces endothelial expression of ICAM-1 and VCAM-1 [4] (Fig. 2).
The RANK-RANKL-OPG signalling axis is considered to be the main regulator of the progression of calcification of the blood vessel wall. In this process, the osteogenic differentiation of VSMCs is of fundamental importance [4]. Fibroblast growth factor 21 (FGF21) fulfils a protective role for the vessels by limiting the osteogenic differentiation of VSMCs. Ecto-5′-nucleotidase (CD73) also plays a protective role for blood vessels, by hydrolysing extracellular adenosine monophosphate (AMP) and, thus, releasing adenosine. Adenosine inhibits the calcification of the blood vessel wall [4]. Adenosine is considered to be a cytoprotective modulator that plays a protective role against tissue damage in various organs. It carries out its biological action by connecting with the G-protein-coupled receptor present on the cell surface, or rather with its subtypes A1, A2a, A2b and A3. There is a link between adenosine triphosphate (ATP)/adenosine metabolism and the RANK-RANKL-OPG signalling axis. It has been confirmed in human osteoprogenitor cells that adenosine inhibits the secretion of OPG [4].
An important connection also exists between OPG and the renin-angiotensin system (RAS), which plays a key role in the development, with age, of unfavourable changes in the VSMC and EC phenotypes and in the pathogenesis of atherosclerosis [4]. The main mediator of RAS, i.e. Ang II, acts directly on the vascular endothelium, leading to its dysfunction, the development of inflammation and the progression of atherosclerosis. Activation of the angiotensin II type 1 (AT1) receptors for Ang II causes, inter alia, an increase in the expression of vascular endothelial growth factors (VEGFs). VEGF-A and VEGF-B increase inflammation and remodelling in blood vessels by activating pro-inflammatory mechanisms and pathological angiogenesis. OPG has been confirmed to strengthen these actions of VEGFs [35]. Atheroma-derived cells harvested during endarterectomy were tested under cell culture conditions. These cells and ECs derived from human dermal microvasculature were grown with and without irbesartan, the blocker of the AT1 receptor for Ang II [36]. It has been confirmed that irbesartan, which blocks the AT1 receptor for Ang II, reduces the concentrations of interleukin-6 (IL-6), interleukin-8 (IL-8) and OPG in both cell types. Moreover, it has been shown that in the cells studied, the blockade of the AT1 receptor for Ang II leads to a reduction in extracellular signal–regulated kinase-1 and kinase-2 (ERK1 and ERK2) expression and restricts their phosphorylation. This phosphorylation of ERK1 and ERK2 is normally induced by RANKL when it is linked to RANK. Similar data was obtained in mice when another blocker of the AT1 receptor for Ang II, losartan, also inhibited RANKL-induced ERK1 and ERK2 phosphorylation [37]. This suggests a convergent action of RANKL and Ang II at the level of ERK1/2 phosphorylation regulation. OPG also has the ability to directly activate ERK1/2 phosphorylation, which is associated with pathological angiogenesis [4, 36, 37].
Moreover, mutual stimulating interactions between OPG and RAS have been confirmed [3, 38,39,40]. In the cells of the vascular walls of both humans and mice, Ang II was confirmed to increase OPG expression, whereas the blockade of the AT1 receptor for Ang II with Irbesartan reduces the expression of OPG in these cells. It has also been shown that, just as Ang II has a dose-dependent effect in increasing OPG expression in vascular wall cells, OPG has a dose-dependent effect in increasing the expression of the AT1 receptor for Ang II [3, 41] (Fig. 2). This is another mechanism by which OPG can promote endothelial dysfunction and the development of atherosclerotic lesions.
Under the influence of Ang II, the expression of the aforementioned VEGF is increased, and this causes overexpression of RANK in ECs and enhances the angiogenic response of these cells to RANKL. RANKL may also have an additional action in maintaining EC integrity and inducing a pro-survival effect on ECs [4]. This action of RANKL on ECs occurs via the PI3-kinase/Akt signalling pathway. OPG inhibits this effect by binding to RANKL, which prevents it from combining with RANK and activating the PI3-kinase/Akt signalling pathway. Blocking PI3-kinase reverses this pro-survival effect of RANKL in relation to ECs [42]. The PI3-kinase/Akt signalling pathway is strongly inhibited by VEGF, whose expression increases under the influence of Ang II [43] (Fig. 2).
More and more experimental data indicates the participation of OPG in promoting the inflammatory process in the blood vessel wall. OPG directly induces the expression of adhesion molecules such as ICAM-1 and VCAM-1 and E-selectin in ECs [4, 5]. Thus, OPG causes increased adherence of leucocytes to the endothelial surface, induces the early stages of endothelial dysfunction and leads to the development of atherosclerotic lesions. At the stage when enhanced recruitment of leucocytes, mainly monocytes, lymphocytes and neutrophils, from the blood takes place and they penetrate deep into the vascular wall, OPG interacts with many pro-inflammatory modulators, such as IL-6, IL-1beta or heparan sulphate proteoglycans (HSPGs) [44]. Under the influence of pro-inflammatory cytokines, the production of OPG in ECs increases significantly, which additionally enhances the expression of adhesion molecules as well as the recruitment and transmigration of monocytes, lymphocytes and neutrophils deep into the vascular wall [4, 5, 17, 45]. Many researchers emphasise the importance of OPG as a chemotactic factor for inflammatory cells infiltrating the vascular wall at an early stage of development of atherosclerotic lesions [4, 17, 44,45,46,47,48]. The participation of pro-inflammatory cytokines such as IL-6 and IL-1beta in this process has become the basis for the development of new therapeutic strategies. Attempts are being made to use monoclonal antibodies that target Il-1beta or the alpha receptor for IL-6 (IL-6Ralpha) — see below [49, 50].
The interaction of the ECs with the activated inflammatory cells and the role of the RANK-RANKL-OPG signalling axis in the formation of atherosclerotic lesions are further emphasised by the fact that both cell types express RANKL. RANKL produces its biological effects in the cells of the vascular wall by combining with RANK in the plasma membrane of these cells. Many of these effects on the ECs have been described above. Additionally, within VSMCs, RANKL, by combining with RANK, increases the activity of matrix metalloproteinases (MMPs). On the other hand, OPG, by binding to RANKL, prevents its association with RANK and inhibits the above-described influence of RANKL on the activity of MMPs in VSMCs [4, 48].
The increase in RANKL expression in ECs and VSMCs under the influence of oxidative stress has also been confirmed. This was confirmed for OxLDL and hydrogen peroxide (H2O2) [51]. The stimulating effect of the incubation of vascular wall cells with OxLDL on RANKL expression in these cells is particularly interesting, and this may reflect changes in the vascular wall of patients with high OxLDL concentrations. Within the vascular wall, RANKL has the ability to activate the degranulation of granulocytes infiltrating the vascular wall [52,53,54]. A protective effect in this respect is induced by FGF21, which reduces the expression of RANKL in the cells of the vascular wall. FGF21 is believed to play a protective role against endothelial damage, which is induced by oxidative stress, and plaque formation, as well as to protect against ischaemic damage to cardiomyocytes [55, 56].
The action of oxidative stress on vascular wall cells is related to the ubiquitin–proteasome system (UPS), which also plays a pathogenetic role in the development of endothelial dysfunction and atherosclerosis. It has also been confirmed that UPS plays an important role in the pathogenesis of congestive HF [57, 58]. In various experimental models of HF, both ischaemic and non-ischaemic, an increase in OPG expression was confirmed. Activation of the RANK-RANKL-OPG axis has been confirmed in a rat post-infarction HF model, and it is believed to be involved in the pathogenesis of HF [59].
The combination of OPG and TRAILMany of OPG’s other mechanisms of action in the CV system may be due to its ability to bind to a ligand other than RANKL, such as TRAIL. TRAIL is a protein belonging to the TNF superfamily and is designated TNFSF 10. TRAIL expression has been confirmed in a wide variety of tissues, including vascular wall cells [3, 6, 60]. In humans, TRAIL is a type II transmembrane protein consisting of 281 amino acids. TRAIL, however, can be cleaved at the stalk domain, and after that, it can bind to two other TRAIL molecules to form a biologically active homotrimer circulating in the blood. TRAIL exerts its biological effect through a binding with its receptors. There are five types of receptors for TRAIL (TRAIL-R). TRAIL-R1 and TRAIL-R2 are agonist receptors and are known as death receptors (DRs). They are type I transmembrane proteins and contain an intracellular death domain (DD) which normally stimulates apoptosis when TRAIL connects to TRAIL-R1 and TRAIL-R2. The remaining receptors for TRAIL that are antagonist receptors are called decoy receptors (DcRs). These include TRAIL-R3, TRAIL-R4 and OPG. TRAIL-R3 and TRAIL-R4 are transmembrane proteins which, however, differ from DR in that they do not have a fully developed intracellular DD. On the other hand OPG is a soluble receptor. When TRAIL is not bound to a DcR, e.g. OPG, then it can bind to TRAIL-R1 and TRAIL-R2 and start a signalling pathway leading to apoptosis. These receptors have a high affinity for TRAIL, which in the form of a homotrimer binds to these receptors, and this results in trimerisation of these receptors. This in turn leads to the formation of a death-inducing signalling complex (DISC), which results in the accumulation of the adapter protein Fas-associated death domain (FADD). FADD is an intermediate link between DR and the pro-domain of the initiator caspase 8. At a later stage within the DISC, dimerisation of caspase 8 molecules occurs, leading to the formation of a mature caspase 8, which activates individual executive caspases, such as caspases 3, 6 and 7. This leads to cell apoptosis [6, 61, 62]. This is the so-called extrinsic apoptosis pathway. In some cell types, this activation of executive caspases must be additionally enhanced by the activation of an additional, internal, mitochondrial apoptotic pathway [6, 62]. In both cases, however, activation of executor kinases is necessary to induce apoptosis in cells sensitive to the pro-apoptotic effects of TRAIL. As for the two transmembrane DcRs for TRAIL (TRAIL-R3 and TRAIL-R4), they differ slightly from one another. TRAIL-R3 is linked to the plasma membrane with the help of a glycosylphosphatidylinositol (GPI) linker and lacks a cytoplasmic fragment. On the other hand, TRAIL-R4 has a shortened DD. Neither of them, however, can send a pro-apoptotic signal or start the apoptotic pathway when linked to TRAIL. Both of these DcRs have the ability to compete with DRs for binding to TRAIL, which, according to some researchers, may constitute a protective mechanism against the pro-apoptotic actions of TRAIL in cells resistant to these TRAIL actions [6]. Such a pro-apoptotic effect of TRAIL is shown in relation to neoplastically transformed cells, cells infected by a virus or the overstimulated cells of the immune system [3, 6, 62]. In studies on TRAIL-knockout mice, it was shown that the absence of TRAIL in these mice was associated with a greater susceptibility to the formation of metastases. This confirms the role of TRAIL in providing the body with protection against the formation and spread of neoplasms [3]. Some researchers have highlighted the fact that the selective induction of apoptosis by TRAIL takes place in transformed, neoplastic or infected cells but not in normal cells. The effect of TRAIL in normal cells, though, is less understood. It has been confirmed, however, that when TRAIL is combined with TRAIL-R1 and TRAIL-R2, the apoptotic pathway is not always triggered. Sometimes pathways responsible for cell survival are activated, such as ERK1/2 or PI3-kinase Akt [3, 62, 63]. It is believed that such differentiated effects of TRAIL in relation to different cells may be caused by the redistribution of individual types of TRAIL receptors or the inhibition of intracellular apoptotic pathways [3]. However, the results of the studies conducted so far are not clear-cut. Despite detailed studies of the relative proportions of individual types of receptors for TRAIL (DR/DcR) in individual types of cells, it is still not possible to fully predict the cell’s response to TRAIL [6,
留言 (0)