
記住我


Gut Pathogens volume 14, Article number: 18 (2022)
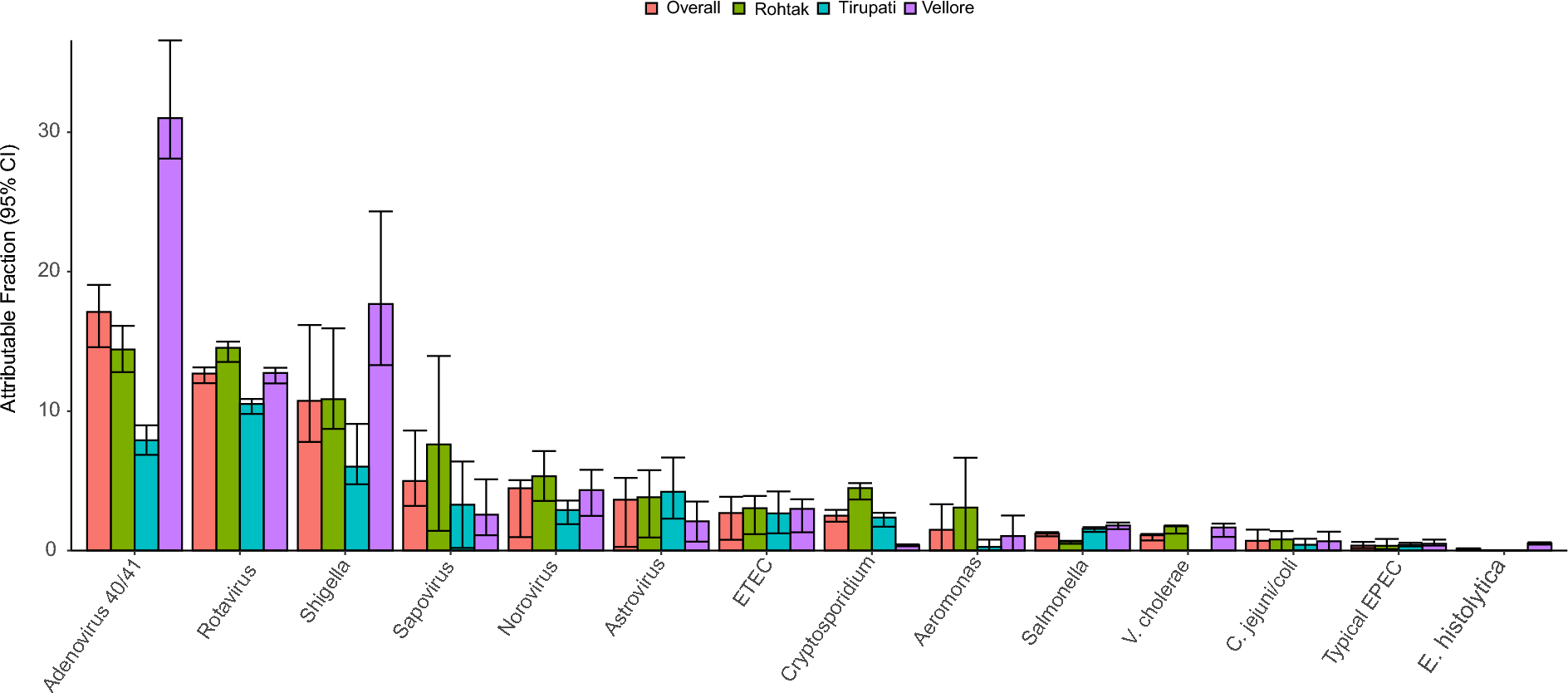
AbstractIn Kuwait, some untreated sewage is discharged into the sea which poses health risks. Therefore, we determined the virulence traits and the phylogenetic groups of E. coli cultured from raw sewage. Sewage was collected once every month for 12 months with culturing of a total of 140 E. coli isolates. E. coli was typed by the methods of Clermont. The five pathotypes of diarrheagenic E. coli (DEC), and extra-intestinal pathogenic E. coli (ExPEC) were detected by specific PCR assays. Four virulence genes which correlate with pathogenicity in animal models, were used for the first time for detection of ExPEC—vat (vacuolating auto-transporter toxin), fyuA (yersiniabactin receptor), chuA (heme-binding protein), and yfcV (major subunit of putative chaperon-usher fimbria). Most E. coli belonged to phylogenetic groups A (65[45%]) and B1 (34[24.3%]). Three (2.1%) isolates were DEC, while 14 (10%) isolates were ExPEC mostly in group B2 (57.1%). A relatively high prevalence of ExPEC in sewage has public health implications.
Sewage is the wastewater consisting of human excreta, wash water, and industrial and agricultural wastes. It is rich in microbes, including bacteria, fungi, parasites, and viruses. The origin of microbes is intestinal (from humans, animals, birds, etc.) and soil. In Kuwait, about 75% of sewage is treated and the remainder is discharged into the sea untreated [1]. Coastal waters are commonly used for recreational purposes in Kuwait.
The discharge of untreated sewage into the coastal waters has health implications for the people who use these waters for recreation. E. coli forms the predominant facultative anaerobic flora of the feces, and hence, it is used as an indicator organism for fecal pollution. Drug-resistant fecal E. coli has been previously cultured from the coastal waters of Kuwait [2]. E. coli can cause both intestinal and extra-intestinal infections [3]. Therefore, we studied E. coli isolates cultured from raw sewage in Kuwait for their virulence properties associated with diarrheal disease and extra-intestinal infections. We also did phylogenetic grouping of the isolates. We defined extra-intestinal pathogenic E. coli (ExPEC) by a set of genes that has not been previously used to screen E. coli from sewage. In addition, we sought all the five pathotypes of diarrheagenic E. coli (DEC) – enteropathogenic E. coli (EPEC), enterotoxigenic E. coli (ETEC), enteroinvasive E. coli (EIEC), shiga toxin-producing E. coli (STEC) and enteroaggregative E. coli (EAEC) [3]—which to our knowledge, has rarely been done, in previous studies of E. coli from sewage.
The Underworlds Project is a collaborative project among MIT-Harvard in the USA, Kuwait Institute for Scientific Research (KISR), Kuwait University, Kuwait Ministry of Public Works, Kuwait Ministry of Health, Kuwait Ministry of Defense, and Kuwait Department of Water and Electricity. This pilot project aims to set up for the State of Kuwait an eventual monitoring system for medicines, phthalates, bioterrorism agents, explosives, antimicrobial resistance genes, bacterial markers of obesity, and infectious disease epidemics (for e.g. due to Salmonella spp., norovirus, rotavirus, influenza virus etc.) by regular analysis of sewage at certain locations. We utilized the E. coli isolates cultured from this project for the present study.
A sewage sample (1000 ml) was collected once every month for 12 months (May 2018 to April 2019) from three sites – Jabriya (latitude: 29.318057, longitude: 48.025805), Zahraa (latitude: 47.99263, longitude: 29.26725) and Hateen (latitude: 29.2842, longitude 48.0184) – with a total of 36 samples. Jabriya is about 7.8 km southeast of Kuwait City, and Zahraa and Hateen are about 11.5 km and 10.8 km respectively south of Kuwait City. Distance between Jabriya and Zahraa is 5.9 km; that between Jabriya and Hateen is 4.1 km; and that between Zahraa and Hateen is 2.1 km. The three sites of sample collection are indicated in Fig. 1. These locations were selected for ease of sample collection. The numbers of people living in these locations at the end of 2018 were: 78,026 (Jabriya), 32,064 (Zahraa) and 21,557 (Hateen) (https://www.citypopulation.de/en/kuwait/admin/). Immediately after the sample was collected, it was transported to KISR in an insulated Styrofoam box with icepacks, for initial processing. After allowing the large particles to settle, a 10.0 ml sample was filtered through a 0.2 μm pore-size membrane (Sartorius). The membrane was immediately transported in cold Styrofoam box to the Enteric Microbiology Laboratory, Faculty of Medicine, Kuwait University, Jabriya, Kuwait, where further processing was done. The membrane was cut into small pieces and vortexed in 1.0 ml of phosphate buffered saline (PBS, pH 7.2). This PBS washing was cultured on m-FC agar (Oxoid) which was incubated at 42 °C for 24 h [4]. Different morphotypes of blue colonies (one to ten) of presumed E. coli were picked and screened for substrate utilization in the API-20E biochemical strip (bioMerieux). Colonies identified as E. coli were stocked in tryptic soy broth (Oxoid) with 15% glycerol at -80˚C for further analysis.
Fig. 1
Locations of Jabriya, Zahraa and Hateen where sewage sampling was done. The map is not drawn to scale
There were some procedures that were common to all PCR assays. These were preparation of template DNA, agarose gel electrophoresis, and detection of amplicons. These are described below. Boiling method was used for DNA extraction. A loopful of E. coli colonies from MacConkey agar was boiled in PCR-grade water (ThermoFisherScientific) in an Eppendorf tube (Eppendorf) for 10 min. After cooling the tube on ice for 5 min, it was centrifuged at 12,000 rpm for 10 min in a Spectrafuge 24D centrifuge (Labnet). The DNA present in the supernatant was used as the template in the PCR assay. The concentration of DNA was measured using a NanoDrop spectrometer (ThermoFisherScientific). The amplification blend was made by mixing the following reagents in a PCR tube (Eppendorf) in a 10 µl reaction volume: 2 µl of master mix (Solis BioDyne), 1 µl of forward primer (10 pmol), 1 µl of reverse primer (10 pmol), 5 µl of PCR-grade H2O, and 1 µl of diluted DNA (25 ng). This PCR mixture was then placed in a thermal cycler (Applied Biosystems) for DNA amplification with the desired cycles. The primer sequences, cycling conditions, and amplicon sizes for various PCR assays are given in Additional file 1: Table S1 at https://github.com/MahdiRedha/GP-AdditionalFile-1Revised14Feb2022/blob/main/GP-AdditionalFile%201Revised14Feb2022.pdf. After amplification, the amplified products were separated by agarose gel electrophoresis. The concentration of the agarose in the gel was 1.0% (for products ˃800 bp), 1.5% (for products 200–800 bp) and 2% (for products < 200 bp). Electrophoresis of the products in the gel incorporated with ethidium bromide was carried out in Tris–Borate-EDTA buffer at a voltage of 100 V for ⁓ 40 min, and the gel was photographed under the UV light to detect amplicons.
Bacterial isolates identified as E. coli by API-20E test were subjected to a confirmatory PCR assay to detect universal stress protein gene (uspA) that is unique for E. coli [5]. Phylogenetic grouping of E. coli was performed by both the triplex and the quadruplex PCR methods [6]. The five categories of DEC were detected by PCR assays for the genes specified in Additional file 1: Table S1. ExPEC was identified by detecting the following genes by PCR assays: vat (vacuolating auto-transporter toxin), fyuA (ferric yersiniabactin uptake A), chuA (coli heme utilization gene A), and yfcV (yersinia fimbrial chaperon V) (Additional file 1: Table S1) by the criteria of Spurbeck et al. [7]; isolates positive for all four genes (vat, fyuA, chuA, yfcV) or the following three gene combinations: fyuA + chuA + yfcV, fyuA + vat + chuA, and fyuA + yfcV + vat were considered as ExPEC.
Of a total of 144 isolates identified as E. coli by API-20E test, 140 isolates were positive by uspA PCR and hence confirmed as E. coli. The number of E. coli cultured each month varied from 3 to 24. E. coli was cultured from Jabriya and Zahraa for all 12 months, and from Hateen for 11 months, except in May 2018 (Additional file 1: Table S2). The proportions of E. coli cultured from the three sites sampled were similar, being 32.8% (46 isolates) in Jabriya, 28.6% (40 isolates) in Zahraa, and 38.6% (54 isolates) in Hateen.
The results of phylogenetic analyses are shown in Additional file 1: Table S2. By triplex PCR grouping, the 140 isolates were classified into group A (68 isolates [48.6%]), group B1 (34 isolates [24. 3%]), group B2 (8 isolates [5.7%]), and group D (27 isolates [19.3%]). As required, isolates from group A and group D were further subjected to quadruplex PCR grouping. By quadruplex PCR, 68 group A isolates were reclassified into group A (65 isolates) and group C (3 isolates); 27 group D isolates were reclassified into group D (15 isolates), group E (4 isolates), and group F (8 isolates). After quadruplex PCR grouping, the rank order of the groups was: A (65 isolates [46.4%]) > B1(34 isolates [24.3%]) > D (15 isolates [10.7%]) > B2 and F (8 isolates [5.7%] each) > E (4 isolates [2.9%]) > C (3 isolates [2.1%]).
The different categories of E. coli were: ExPEC (14 isolates [10%]), DEC (3 isolates [2.1%]) and nonpathogenic E. coli (123 isolates [87.9%]). Further characterization is as below. Of the three DEC, two were atypical EPEC (eae+bfp−) (one cultured from June 2018 sample from Jabriya [isolate no. J28], and the other cultured from November 2018 sample from Hateen [isolate no. H164]), and one was an ltA + ETEC (cultured from July 2018 sample from Hateen [isolate no. H51]). The distribution of the genes that defines 14 isolates as ExPEC is shown in Table 1. The isolates had all four genes and two different combinations of three genes. The distribution of ExPEC according to date and site of collection is shown in Additional file 1: Table S3. ExPEC was isolated from all three sites except for the months of May, June, September, November, and December 2018 when it was not isolated from any site. The rank order of distribution of phylotypes in ExPEC was B2 (8 isolates [57.1%] > F (5 isolates [35.7%] > B1 (1 isolate [7.1%]). Nonpathogenic E. coli (not belonging to DEC or ExPEC), was in group A (65 isolates [52.9%]), B1 (32 isolates [26.0%]), D (14 isolates [11.4%]), E (4 isolates [3.3%]), C and F (3 isolates each [2.4%]), and B2 (2 isolates [1.6%]).
Table 1 Distribution of genes in E. coli isolates which define ExPEC, and phylotypes of ExPECE. coli bacteria are heterogeneous and constitute commensals, intestinal pathogens, and extra-intestinal pathogens. Grouping of E. coli can be done using molecular methods. One such method is the PCR phylogenetic typing done in our study. This typing method was developed by Clermont et al. [6]. They initially did the typing with a triplex PCR method using three genes (chuA, yjaA, and TspE4.C2), and assigned E. coli to four groups: A, B1, B2, and D [6]. When they compared the typing results with those of the multi-locus sequence typing, it was found that assigning some isolates to groups A and D was not accurate. Therefore, they developed a modified method, a quadruplex PCR method [6]. Quadruplex method uses modified primers targeting the same three genes (chuA, yjaA, and TspE4.C2), thus eliminating some problems associated with mismatches of the primers [6]. It also uses primers to target an additional gene, arpA. By this quadruplex PCR, some isolates of phylogroups A and D were further divided into groups C, E and F in our study. It was reported that quadruplex PCR also identifies cryptic clades I to V, which were not found in our study. Cryptic E. coli clades are extremely rare in human samples. They are mainly found in feces of animals, non-human mammals, and birds [8]. The proportion of reclassified E. coli phylogroups A and D in our study, was as predicted by Clermont et al. [6]. It has been shown that the PCR phylogenetic grouping has similar discriminatory power as enterobacterial repetitive intergenic consensus (ERIC) PCR and the laborious pulsed-field gel electrophoresis (PFGE) [9].
Only three of the 140 E. coli isolates belonged to DEC (2.1%). This low prevalence of DEC is in line with a study in children in Kuwait [10]. ExPEC pathotype is the causative agent of urinary tract infection, bacteremia/septicemia, and meningitis. It harbors the highest number of virulence factors that include adhesins, toxins, siderophores and hemolysins [11]. In one study [9], after final treatment of the sewage, the effluent had E. coli that harbored virulence factors specific for ExPEC suggesting that ExPEC strains can survive sewage treatments.
ExPEC strains do not have a defined set of genes as in the case of DEC. However, studies have shown that there are some genes that are commonly found in ExPEC pathotype in high frequencies, differentiating ExPEC from DEC and commensals. Spurbeck et al. [7] showed that four genes—vat, fyuA, chuA, and yfcV—could be used to identify uropathogenic E. coli (UPEC) isolates. By their criteria, isolates positive for all four genes (vat, fyuA, chuA, yfcV) or the following combinations of genes—fyuA + chuA + yfcV, fyuA + vat + chuA, and fyuA + yfcV + vat—were ExPEC. Isolates with these genes were found to have more virulence determinants and could colonize the bladder more effectively. The study of Johnson et al. [12] found that the following genes—papA/papC, sfa/focDE, afa/draBC, iutA, and kpsMT II—were highly associated with ExPEC isolates. Screening a collection of E. coli isolates by the tests of Spurbeck et al. [7] and Johnson et al. [12] showed that the isolates predicted as UPEC or ExPEC had most additional virulence genes [7]. Another study done by Johnson et al. [13] suggested that the tests of Spurbeck et al. [7] were valid for fecal isolates. Thus, it appeared that we could either use the criteria of Spurbeck et al. [7] or those of Johnson et al. [12] to define ExPEC. We chose the criteria of Spurbeck et al. [7] in our study. The four genes used in our study to define ExPEC appeared to correlate with virulence in a mouse model [7]. Thus, our criteria for detecting ExPEC have a high reliability compared to criteria used in other studies. To our knowledge, ours is the first study, where these four genes were used to identify ExPEC from sewage.
In previous studies, most of the ExPEC belonged to groups B2 and D, while those E. coli that belonged to group A or B1 were recognized as commensals [14, 15]. It was also demonstrated that isolates from group B2 were more virulent than the isolates from the other groups [15]. In our study too, more than half of the ExPEC isolates belonged to group B2.
Some studies have suggested that strains belonging to groups A and B1 may also harbor virulence traits that enable them to survive and proliferate in the harsh environments outside the gastrointestinal tract [16]. Some of these strains may cause intestinal and extra-intestinal illnesses as they possess the required virulence genes. Thus, the two groups, A and B1, should not be dismissed as non-pathogenic [10, 17].
Probably, a limitation of our study is its complete reliance on PCR methods for detection of virulence genes. The four genes that were used for the detection of ExPEC were based on the expression of these genes in in vivo models by the original investigators [7]. We are not aware of any studies that showed a lack of expression or non-functionality of these genes due to mutation. Even so, a future study should look at the expression of these virulence genes that define ExPEC, in isolates from sewage. Therefore, the observations of our study may be considered as preliminary.
The characteristics of E. coli in raw sewage in Kuwait reflects E. coli population in the colonic flora of the Kuwaiti population. The finding of a relatively high prevalence of ExPEC in raw sewage in Kuwait is of a public health concern for the use of coastal waters for recreation. A future study that examines the carriage of ExPEC virulence genes by E. coli isolates recovered from contaminated coastal waters is warranted. The dilution of the organisms by the sea water as a mitigating factor must also be taken into consideration.
Availability of data and materialsAll data generated or analyzed during this study are included in this published article [and its supplementary information file].
AbbreviationsDEC:Diarrheagenic E. coli
EAEC:Enteroaggregative E. coli
STEC:Shiga toxin-producing E. coli
EIEC:Enteroinvasive E. coli
EPEC:Enteropathogenic E. coli
ETEC:Enterotoxigenic E. coli
ExPEC:Extra-intestinal pathogenic E. coli
UPEC:Uropathogenic E. coli
ReferencesAleisa E, Alshayji K. Analysis on reclamation and reuse of wastewater in Kuwait. J Eng Res. 2019;7:1–13.
Alsarawi HA, Jha AN, Baker-Austin C, Alsarawi MA, Lyons BP. Baseline screening for the presence of antimicrobial resistance in Escherichia coli isolated from Kuwait’s marine environment. Marine Poll Bull. 2018;129:893–8.
Kaper JB, Nataro JP, Mobley HL. Pathogenic Escherichia coli. Nat Rev Microbiol. 2004;2:123–40.
Park JE, Ahn TS, Lee HJ, Lee YO. Comparison of total and fecal coliforms as faecal indicator in eutrophicated surface water. Water Sci Technol. 2006;54:185–90.
Chen J, Griffiths MW. PCR differentiation of Escherichia coli from other Gram-negative bacteria using primers derived from the nucleotide sequences flanking the gene encoding the universal stress protein. Lett Appl Microbiol. 1998;27:369–71.
Clermont O, Christenson JK, Denamur E, Gordon DM. The Clermont Escherichia coli phylo-typing method revisited: Improvement of specificity and detection of new phylo-groups. Environ Microbiol Rep. 2013;5:58–65.
Spurbeck RR, Dinh PC Jr, Walk ST, Stapleton AE, Hooton TM, Nolan LK, et al. Escherichia coli isolates that carry vat, fyuA, chuA, and yfcV efficiently colonize the urinary tract. Infect Immun. 2012;80:4115–22.
Clermont O, Gordon DM, Brisse S, Walk ST, Denamur E. Characterization of the cryptic Escherichia lineages: rapid identification and prevalence. Environ Microbiol. 2011;13:2468–77.
Anastasi EM, Matthews B, Stratton HM, Katouli M. Pathogenic Escherichia coli found in sewage treatment plants and environmental waters. Appl Environ Microbiol. 2012;78:5536–41.
Albert MJ, Rotimi VO, Dhar R, Silpikurian S, Pacsa AS, Molla AM, et al. Diarrhoeagenic Escherichia coli are not a significant cause of diarrhoea in hospitalised children in Kuwait. BMC Microbiol. 2009;9:62.
Mobley H, Donnenberg M, Hagan E. Uropathogenic Escherichia coli. EcoSal Plus. 2009. https://doi.org/10.1128/ecosalplus.8.6.1.3.
Johnson JR, Murray AC, Gajewski A, Sullivan M, Snippes P, Kuskowski MA, et al. Isolation and molecular characterization of nalidixic acid-resistant extraintestinal pathogenic Escherichia coli from retail chicken products. Antimicrob Agents Chemother. 2003;47:2161–8.
Johnson JR, Johnston BD, Porter S, Thuras P, Aziz M, Price LB. Accessory traits and phylogenetic background predict Escherichia coli extraintestinal virulence better than does ecological source. J Infect Dis. 2019;219:121–32.
Duriez P, Clermont O, Bonacorsi S, Bingen E, Chaventre A, Elion J, et al. Commensal Escherichia coli isolates are phylogenetically distributed among geographically distinct human populations. Microbiology. 2001;147:1671–6.
Micenková L, Bosák J, Vrba M, Ševčíková A, Šmajs D. Human extraintestinal pathogenic Escherichia coli strains differ in prevalence of virulence factors, phylogroups, and bacteriocin determinants. BMC Microbiol. 2016;16:218.
留言 (0)