
記住我




Expansion of the blood vessel network is prerequisite not only for development but also in the adult organism during tissue growth, regeneration, or wound healing. While insufficient blood vessel formation prevents regeneration of ischemic tissue, excessive and deregulated angiogenesis is implicated in several ocular diseases and cancer (Potente et al, 2011). In the last years, it became clear that angiogenesis is not only coordinated by growth factors and signaling transduction pathways but is also dependent on adaption in cellular metabolism (Eelen et al, 2018). For example, sprouting of new vessel branches is dependent on increased glycolysis (De Bock et al, 2013; Schoors et al, 2014), whereas fatty acid beta-oxidation (FAO) is key for biomass generation and endothelial cell (EC) proliferation (Schoors et al, 2015). Also, lymphatic EC proliferation, migration, and sprouting rely on FAO (Wong et al, 2017). A recent study reported that lymphatic ECs can not only oxidize fatty acids but also ketone bodies and that this promotes formation of new lymphatic vessels in mice (Garcia-Caballero et al, 2019; Puchalska & Crawford, 2019). However, it has not yet been reported whether also vascular ECs can take up and oxidize ketone bodies and if this would affect their angiogenic properties.
Ketone body metabolism enables mammals to endure periods of carbohydrate restriction. During starvation, intensive endurance sports or consumption of carbohydrate-restrictive diets, glycogen stores get depleted and insulin levels decline. This stimulates lipolysis of triacylglycerides in adipocytes and release of free fatty acids and glycerol into the bloodstream. Free fatty acids are taken up by non-adipocytes and broken down into acetyl-CoA moieties via FAO to fuel the tricarboxylic acid (TCA) cycle, thereby contributing to energy production (Puchalska & Crawford, 2017). In the liver, however, TCA cycle intermediates are shunt into gluconeogenesis in situations of glucose deprivation. This results in a surplus of acetyl-CoA in hepatic mitochondria whenever FAO-derived acetyl-CoA exceeds flux through the TCA cycle. Under these circumstances, acetyl-CoA molecules are enzymatically converted into the ketone bodies acetoacetate and β-hydroxybutyrate (Puchalska & Crawford, 2021). In the healthy, fed state serum ketone body (acetoacetate and β-hydroxybutyrate) concentrations in humans are low (50–200 µM). Physiological ketosis, for example, during starvation increases serum ketone body concentrations to around 1–2 mM, whereas pathological ketosis, as observed in untreated type 1 diabetes patients, is characterized by very high serum ketone body concentrations up to 20 mM (Robinson & Williamson, 1980).
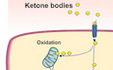
Newly synthesized ketone bodies are released from hepatocytes into the bloodstream and function as energy-rich alternative metabolic fuel in particular for heart and skeletal muscle. The rate of ketone body oxidation in extrahepatic organs increases proportionally to serum ketone body concentrations. In myocytes, D-β-hydroxybutyrate dehydrogenase (BDH1) catalyzes the oxidation of β-hydroxybutyrate, the predominant ketone body molecule in blood, to acetoacetate. Acetoacetate is then transferred onto CoA through the activity of succinyl-CoA:3-oxoacid-CoA transferase (SCOT, encoded by 3-oxoacid CoA-transferase 1 (Oxct1)). Acetoacetyl-CoA can then catalyzed by mitochondrial thiolase into acetyl-CoA fueling the TCA cycle for energy production (Cotter et al, 2013).
Notably, ketone bodies and other circulating nutrients need to pass the endothelial barrier, the inner surface of blood vessels, before being taken up by tissue cells. Whereas the sinusoidal endothelium in the liver allows free diffusion, the continuous endothelium in, for example, muscle, lung, or brain requires transcellular transport (Augustin & Koh, 2017). This raises the question whether ECs function to transport ketone bodies from blood toward tissue cells only or whether they are also able to use ketone bodies themselves. Based upon the outstanding importance of cellular metabolism for controlling angiogenesis (Eelen et al, 2018), this study aimed at analyzing ketone body metabolism in vascular ECs. The study focused on cardiac ECs, as the heart is one of the main consumers of ketone bodies during physiological ketosis.
Results Cardiac ECs oxidize ketone bodies via the TCA cycleOxidation of ketone bodies (Fig 1A) requires the key enzyme SCOT. Heart, skeletal muscle, and brain are the main consumers of ketone bodies (Puchalska & Crawford, 2017) and SCOT protein was abundantly present in tissue lysates of these organs obtained from adult C57Bl/6J mice, as expected (Fig 1B and C). We also detected weak SCOT expression in adipose tissue, whereas it was not detectable in lung and liver. Expression of the oxidoreductase BDH1, which reversibly converts D-β-hydroxybutyrate into acetoacetate, showed a variable organ expression pattern with highest expression in the liver, the organ that is responsible for ketone body production (Fig 1B and D). In the heart, cardiomyocytes oxidize ketone bodies, but whether other cell types in the heart also oxidize ketones is unknown (Schugar et al, 2014; Horton et al, 2019). Indeed, we detected high SCOT and BDH1 expression levels in freshly isolated murine cardiomyocytes. Surprisingly however, we also observed expression of both proteins in the non-cardiomyocyte fraction, in particular in CD31+ vascular ECs (Fig 1G).

Figure 1. Key enzymes of ketone body oxidation are expressed in cardiac endothelial cells
A. Oxidation of ketone bodies requires the enzymes SCOT and BDH1 and yields acetyl-CoA which enters the tricarboxylic acid (TCA) cycle. B. Immunoblot of SCOT, BDH1, and the loading control VCP across various organs obtained from adult C57Bl/6J mice. C, D. Quantification of SCOT and BDH1 expression levels relative to VCP in various murine organs. E. Immunoblot of SCOT, BDH1, CD31, and the loading control VCP in murine endothelial cells isolated from heart, muscle, lung, and adipose tissue. F. Immunoblot of SCOT, BDH1, CD31, and the loading control VCP in human cardiac ECs, HUVECs, and adipose tissue ECs. G. Immunoblot of SCOT, BDH1, CD31, and VCP in cardiac endothelial cells (EC) and non-endothelial cell fraction (non-EC) isolated from adult C57Bl/6J mice. H. Immunoblot of SCOT, BDH1, and β-actin in immortalized murine cardiac endothelial cells (MCEC). I, J. Ketone body concentration in cell culture medium of MCEC treated with 2 mM D-β-hydroxybutyrate (β-OHB) or 2 mM acetoacetate (AcAc) after 24 h. Data are presented as mean ± SD. n = 3; *P < 0.05; ***P < 0.001 unpaired Student’s t-test.Source data are available online for this figure.
Endothelial SCOT and BDH1 protein expression was not only restricted to cardiac ECs but also detectable in ECs derived from skeletal muscle, lung, and adipose tissue of adult mice (Fig 1E). Also, ECs derived from human umbilical cord vein (HUVEC), heart, and adipose tissue expressed SCOT and BDH1 protein (Fig 1F). Analysis of single-cell RNA sequencing data from isolated ECs of multiple murine organs (Kalucka et al, 2020) revealed that Oxct1 mRNA (encoding SCOT protein) gets transcribed in ECs of all organs analyzed. In the heart, Oxct1 mRNA expression was uniformly distributed between arterial, capillary, and venous ECs (Appendix Fig S1). To test for potential upstream regulators of Oxct1 and Bdh1, immortalized murine cardiac ECs (MCEC) as well as primary human cardiac microvascular ECs and HUVECs were treated with inflammatory cytokines (IL1β, IL6, TNFα), pro-angiogenic VEGF-A, cultured under normoxic versus hypoxic conditions and starved versus FCS-treated conditions. However, these experiments did not lead to the identification of an upstream regulatory factor (Appendix Fig S2).
To test whether expression of SCOT and BDH1 would allow vascular ECs to metabolize ketone bodies, we first analyzed whether MCEC are able to take up ketone bodies from the culture medium. These cells express the ketolytic enzyme SCOT as well as BDH1 (Fig 1H). When MCEC were cultured in the presence of 2 mM acetoacetate or 2 mM D-β-hydroxybutyrate, there was a marked decrease of ketone body concentration 24 h later in the culture medium (Fig 1I and J). This indicates that MCEC take up ketone bodies.
Interestingly, targeted metabolite profiling also showed that treatment of MCEC with both acetoacetate or D-β-hydroxybutyrate increased the absolute concentration of several tricarboxylic acid (TCA) cycle intermediates such as citrate (Fig 2A and B). To investigate the ability of MCEC to metabolize ketone bodies, isotope tracing untargeted metabolomics using ketone bodies was performed. MCEC that were treated with uniformly labeled [U-13C4] acetoacetate or [U-13C4] D-β-hydroxybutyrate for 24 h had incorporated the 13C isotope into numerous metabolites indicating that cardiac ECs do not only take up ketone bodies but also metabolize them. Most importantly, 13C incorporation was observed in TCA cycle intermediates, amino acids and putative purines and lipids (Fig 2C and D). The 13C incorporation rate was almost always higher in the presence of acetoacetate compared to D-β-hydroxybutyrate. This is most likely due to the fact that D-β-hydroxybutyrate oxidation requires an additional enzymatic oxidation step into acetoacetate by BDH1 (Fig 1A). Together, these data indicate that cardiac ECs take up ketone bodies to feed the TCA cycle and to generate biomass.

Figure 2. Ketone body treatment increases TCA cycle intermediates in MCEC
A, B. Murine cardiac endothelial cells (MCEC) were treated with 2 mM D-β-hydroxybutyrate (β-OHB) or 2 mM acetoacetate (AcAc) for 24 h. Quantification of concentrations of TCA cycle intermediates compared to control treatment. C. Scheme of ketone body oxidation and the tricarboxylic acid (TCA) cycle. MCEC were incubated with 1 mM 13C4-β-hydroxybutyrate or 1 mM 13C4-acetoacetate for 24 h or the same concentration of unlabeled ketone bodies ([12C] isotope). The [13C]-labeled fractions of the TCA cycle intermediates citrate, α-ketoglutarate and malate (marked with blue star) were quantified with an untargeted metabolomics approach indicating that 13C carbon from ketone bodies was incorporated into TCA cycle intermediates. D. Quantifications of [13C]-labeled fractions of amino acids (asparagine, l-aspartate, l-glutamate, l-proline), putative lipid species (nervonic acid, 1-palmitoylglycerol, ceramide (d18:1/16:0)) and uridine diphosphate (UDP) species (uridine monophosphate, uridine 5'-diphosphate, UDP-N-acetylglucosamine) in MCEC treated with 1 mM 13C4-β-hydroxybutyrate or 1 mM 13C4-acetoacetate for 24 h or the same concentration of unlabeled ketone bodies ([12C] isotope). Data are presented as mean ± SD. n ≥ 4; ns, not significant; *P < 0.05; **P < 0.01; ***P < 0.001 unpaired Student’s t-test. Ketone bodies are a source for mitochondrial respiration in cultured ECsTo test whether ketone bodies can also be oxidized by cultured ECs to feed to mitochondrial electron transfer chain and ATP production, we employed Seahorse technology (XF Cell Mito Stress Kit). This revealed that the addition of ketone bodies to the culture medium of serum-starved MCEC allows for an increase in maximal mitochondrial respiration and ATP production (Fig EV1A–H).
Click here to expand this figure.
Figure EV1. Effects of ketone bodies on mitochondrial respiration
Mitochondrial function of MCECs stimulated with ketone bodies was characterized using the Seahorse Bioanalyzer by measuring the rate of oxygen consumption (OCR) following sequential additions of oligomycin, FCCP and antimycin/rotenone (A) to the cells to determine non-mitochondrial oxygen consumption (B), basal respiration (C), maximal respiration (D), proton leakage (E), ATP production (F), spare respiratory capacity (G), and the bioenergetic health index (H), respectively. A–H. MCECs were starved for 1 h before addition of 2 mM R-β-hydroxybutyrate (βOHB) compared to H2O and acetoacetate (AcAc) compared to ethanol for 24 h. I–L. MCECs were cultured in basal medium before addition of ketone bodies (2 mM) and solvent control. (I, J) Cellular ATP contents after 12 and 24 h. (K, L) Targeted metabolomics using UPLC to determine cellular nucleotides upon treatment with ketone bodies for 24 h. Data are presented as mean ± SD. One-way ANOVA using nonparametric (Kruskal–Wallis) test. ****P < 0.0001, ***P < 0.001, **P < 0.01, *P < 0.05, NS > 0.05.When MCEC were cultured in full medium under basal conditions, further addition of ketone bodies did not increase the amount of ATP (Fig EV1I and J). Also, the amounts of other nucleotides were not changed by addition of ketone bodies (Fig EV1K and L). As ATP cannot be sufficiently stored by cells, this finding was not surprising. In summary, the experiments shown so far revealed that ECs can uptake and oxidize ketone bodies to either produce ATP (e.g., during starvation) or biomass.
Ketone bodies promote angiogenic capacity of cultured cardiac ECsAs biomass production is key for EC growth and cell division (Eelen et al, 2018), we investigated whether the presence of ketone bodies would allow cardiac ECs to proliferate faster. In the presence of D-β-hydroxybutyrate or acetoacetate in the culture media, MCEC showed higher levels of BrdU incorporation as a marker of DNA synthesis (Fig 3A and B). When cells were cultured on gold microelectrodes, elevated electrical impedance was observed, indicating increased cell proliferation rates in response to ketone body treatment (Fig 3C and D). Consistently, treatment with ketone bodies (0.1–10 mM) increased the number of MCEC upon 24, 48, and 72 h of culture compared to a solvent control (Fig EV2A–F). In addition, MCEC exposed to D-β-hydroxybutyrate or acetoacetate migrated faster in a Boyden chamber assay when compared to control treatment (Fig 3E and F).

Figure 3. Ketone body treatment promotes endothelial cell proliferation, migration, and sprouting capacity
A, B. Murine cardiac endothelial cells (MCEC) were treated with different concentrations of D-β-hydroxybutyrate (βOHB) or acetoacetate (AcAc) for 24 h. Relative BrdU absorbance was quantified compared to control treatment (H2O for βOHB and EtOH for AcAc). C, D. MCEC were treated with 10 mM D-β-hydroxybutyrate (βOHB) or 10 mM acetoacetate (AcAc) for 24 h. Electrical impedance was measured compared to control treatment (dashed line; H2O for βOHB and EtOH for AcAc). E, F. MCEC were seeded onto Boyden chambers and treated with (E) 10 mM D-β-hydroxybutyrate (βOHB) or (F) 10 mM acetoacetate (AcAc) for 24 h. Electrical impedance was measured in the lower chamber compared to control treatment (dashed line; H2O for βOHB and EtOH for AcAc). G, H. Representative images of MCEC spheroids treated with H2O, 30 mM R-β-hydroxybutyrate (βOHB), ethanol (EtOH) or 30 mM acetoacetate (AcAc) for 72 h; scale bar: 50 µm. I–M. Angiogenic capacity of MCEC in response to ketone body treatment was analyzed using a spheroid-based sprouting assay. Spheroids were treated with media containing 30 mM D-β-hydroxybutyrate (βOHB), 30 mM acetoacetate (AcAc), 10 mM acetate, 1 mM octanoate, 1 mM butyrate, 30 mM L-β-hydroxybutyrate (L- βOHB), 1 mM niacin or the respective controls (final concentration of reagents is diluted to approximately 10%) and analyzed after 48 h. The (I, L) average number of sprouts per spheroid, the (J, M) accumulated total sprout length and (K) average sprout length were quantified. Data are presented as mean ± SD. n ≥ 3; *P < 0.05; **P < 0.01; ***P < 0.001 unpaired Student’s t-test. Click here to expand this figure.
Figure EV2. Effects of ketone body supplementation on sprouting potential
A–C. Cell counts of MCECs treated with 0.1, 1 or 10 mM R-β-hydroxybutyrate (βOHB) for 24, 48, or 72 h compared to treatment with H2O (control). D–F. Cell counts of MCECs treated with 0.1, 1 or 10 mM acetoacetate (AcAc) for 24, 48, or 72 h compared to treatment with ethanol (control). G. Representative images of HUVEC spheroids treated with H2O (control), recombinant VEGF-A165, 30 mM R-β-hydroxybutyrate (βOHB) for 72 h; scale bar: 50 μm. H. Quantification of the accumulated total sprout length per spheroid. n = 4. I. Relative absorbance of BrdU incorporated into DNA of HUVEC treated with 1, 4 or 10 mM R-β-hydroxybutyrate (βOHB), 0.1, 1 or 10 mM acetoacetate (AcAc) or a combination of both for 24 h. Data are presented as mean ± SD. n ≥ 2. One-way ANOVA; *P < 0.05; **P < 0.01; ***P < 0.001.EC proliferation and migration are prerequisites for angiogenesis, the formation of new blood vessels from pre-existing ones (Potente et al, 2011). In a three-dimensional spheroid-based angiogenesis assay in collagen, treatment with D-β-hydroxybutyrate or acetoacetate resulted in increased capillary sprout formation (Fig 3G and H), as quantified by the numbers of sprouts and the cumulative and average sprout length (Fig 3I–K). Similar results were also obtained with human umbilical vein ECs (HUVEC), indicating that this effect is not restricted to cardiac-specific ECs (Fig EV2G–I).
To test whether this effect is specific for D-β-hydroxybutyrate and acetoacetate, MCEC spheroids were cultured in the presence of other substrates which can feed into the TCA cycle such as acetate, the short-chain fatty acid butyrate, or the medium-chain fatty acid octanoate. However, these did not promote the angiogenic potential (Fig 3L and M). To rule out that the observed pro-angiogenic effect of ketone bodies on cardiac ECs is based on oxidation-independent mechanisms, we performed a series of experiments. First, MCEC were treated with ketone body enantiomer L-β-hydroxybutyrate, that is not a substrate of BDH1 and can therefore not be oxidized. Notably, only D-β-hydroxybutyrate (Fig 3I–K), but not L-β-hydroxybutyrate (Fig 3L–M) promoted tube formation capacity of MCEC. Secondly, we addressed the possibility whether D-β-hydroxybutyrate would exert its functions by inhibiting class I histone deacetylases (HDAC) as this has been shown by treating mice with high doses (Shimazu et al, 2013). In ECs, there are contradictory reports about the efficiency of D-β-hydroxybutyrate acting as a HDAC inhibitor (Chriett et al, 2019; Li et al, 2021). To rule out the possibility that increased angiogenic potential of MCEC would merely be the consequence of HDAC inhibition, MCEC were treated with the well-established HDAC inhibitor butyrate (Shimazu et al, 2013). In contrast to D-β-hydroxybutyrate, butyrate treatment had no effect on the tube formation capacity, indicating that the observed effects are unlikely to be mediated through histone acetylation (Fig 3L and M). Lastly, D-β-hydroxybutyrate has been shown to act as a ligand for hydroxycarboxylic acid receptor-2 (HCA2, also known as niacin receptor-1 (NICAR1) or GPR109A) (Graff et al, 2016). We aimed at ruling out that the proangiogenic effect of D-β-hydroxybutyrate would be mediated through stimulation of HCA2. Therefore, MCEC were treated with niacin, an alternative ligand for HCA2. There were no differences in the angiogenic potential of MCEC in response to niacin when compared to control treatment, ruling out that niacin receptor signaling plays a substantial role in the observed phenotype (Fig 3L and M).
SCOT is necessary for the pro-angiogenic effects of ketone bodiesThe data presented so far indicate that oxidation of ketone bodies is subsequently promoting cell proliferation and tube formation in ECs. To further prove this, we generated SCOT-deficient MCEC lines using CRISPR/Cas9 technology. This resulted in a complete loss of SCOT protein expression in MCEC (Fig 4A), which did not interfere with cell viability. However, SCOT-deficient MCEC did not show an increased angiogenic potential in response to ketone body treatment in a spheroid-based sprouting assay compared to control cells transfected with a non-targeting plasmid construct (Fig 4B and C). In summary, the data reported thus far indicate that the oxidation of β-hydroxybutyrate and acetoacetate is both mandatory and responsible for the increased proliferation and migration rates as well as the higher sprouting ability of ECs in response to elevated ketone body concentrations in the supernatant.

Figure 4. Ketone body treatment does not increase sprouting capacity in SCOT-deficient cardiac endothelial cells
A. SCOT-deficient murine cardiac endothelial cells were generated using the CRISPR/Cas9 technology by targeting the SCOT-encoding gene Oxct1. A non-targeting control construct was used to generate the non-targeting control cells (nt ctrl). Immunoblot of SCOT and β-actin in non-targeting control cells (nt ctrl) and Oxct1 knockout cells (Oxct1 ko). B, C. Angiogenic capacity of Oxct1 knockout cells in response to ketone body treatment was analyzed using a spheroid-based sprouting assay. Spheroids were treated with media containing 30 mM D-β-hydroxybutyrate (βOHB), 30 mM acetoacetate (AcAc) or the respective controls (final concentration of reagents is diluted to approximately 10%) for 48 h. The average number of sprouts per spheroid and the accumulated sprout length was quantified. Data are presented as mean ± SD. n = 4; *P < 0.05; **P < 0.01; unpaired Student’s t-test.Source data are available online for this figure.
Ketogenic diet alters gene expression of cardiac ECs in miceWe next investigated the effects of elevated serum ketone body concentrations on cardiac ECs in C57Bl/6J mice. As expected, administration of a high-fat, very low-carbohydrate ketogenic diet increased the ketone body concentration in blood (Fig 5A). To gain insight into the changes in gene expression caused by elevation of serum ketone body concentrations, cardiac CD31+ ECs were isolated (Fig EV3A and B) from mice that were fed a ketogenic diet or control diet and subjected to RNA-sequencing. This revealed that feeding a ketogenic diet for 2 weeks changed the expression of around 150 genes in cardiac ECs when compared to cells of littermates kept on a control diet (P-value ≤ 0.01, log2 fold change ≤ −0.5 or ≥ 0.5) (Fig 5B–D, Dataset EV1). Ketogenic diet led to no changes in BDH1 expression. However, expression of Oxct1 (SCOT) was significantly lower at 3 and 7 days after starting the diet, but no longer at 14 days (Fig EV3C). We performed a gene set enrichment analysis (GSEA) using the annotated hallmark gene sets from the molecular signature database (MSigDB) and observed a significant enrichment of genes involved in fatty acid metabolism (Fig 5E and F). The mitochondrial pyruvate dehydrogenase lipoamide kinase isozyme 4 (Pdk4) was significantly upregulated in cardiac ECs (Figs 5D and EV3D). Pdk4 inhibits the pyruvate dehydrogenase complex, thereby regulating the entry of glycolysis-derived acetyl-CoA into the TCA cycle and is also known to be upregulated in whole heart tissue lysates in response to ketogenic diet (Wentz et al, 2010). The upregulation of Pdk4 and Hmgcs2 expression in cardiac ECs during ketogenic diet (Fig 5D) was confirmed in a second cohort of mice (Fig EV3D).

Figure 5. Ketogenic diet alters gene expression in cardiac endothelial cells towards a more proliferative signature
A. Ketone body concentrations were measured in serum of adult C57Bl/6J mice that were fed a ketogenic diet for the indicated time periods. B. RNA sequencing was performed on isolated cardiac endothelial cells of male C57Bl/6J mice that were fed a ketogenic diet or control diet for 2 weeks. Volcano plot showing differentially expressed genes as genes with a log2 fold change ≤ −0.5 or ≥ 0.5 (vertical dashed lines) and P ≤ 0.01 (horizontal dashed line). C, D. Heatmaps showing expression levels of genes involved in KEGG pathways (C) fatty acid degradation and (D) ketone body metabolism in cardiac endothelial cells isolated from male C57Bl/6J mice that were fed a ketogenic diet or control diet for 2 weeks. E. Gene set enrichment analysis of hallmark (GSEA gene set h, blue) and gene ontology (GO)-derived (GSEA gene set c5, green) gene sets enriched in cardiac endothelial cells of C57Bl/6J mice kept on a ketogenic diet compared to littermate animals kept on control diet. F, G. Enrichment plots of the hallmark gene sets fatty acid metabolism and G2M checkpoint comparing expression pattern of cardiac endothelial cells isolated from C57Bl/6J mice kept on a ketogenic diet compared to littermate animals kept on control diet. Data are presented as mean ± SD. n ≥ 4; ***P < 0.001; unpaired Student’s t-test. Click here to expand this figure.
Figure EV3. Cardiac endothelial cells isolated from mice fed a ketogenic diet
Relative mRNA levels of cardiomyocyte, fibroblast, and smooth muscle cell marker genes in anti-CD31-isolated cells compared to levels in total heart lysates of C57Bl/6J mice. Flow cytometry analysis of anti-CD31-isolated cells from hearts of C57Bl/6J mice using the endothelial cell marker CD31 and the immune cell marker CD45. Relative fold changes of Oxct1 (Scot) and Bdh1 expression levels in mice fed a ketogenic diet relative to a control diet were obtained from RNAseq analyses. Relative mRNA levels of cardiac ECs isolated from mice fed a ketogenic or control diet confirming increased levels of pro-proliferative genes Hmgcs2 and Pdk4. Heat-map showing expression levels of genes involved in regulation of cell cycle (KEGG). Data are presented as mean ± SD. n ≥ 3. Two-tailed unpaired Student’s t-test; **P < 0.01; ***P < 0.001.Further GSEA showed an enrichment of genes involved in processes related to cell cycle progression and cell proliferation such as gap 2-mitosis (G2/M) checkpoint and E2F targets (hallmark gene sets) as well as sister chromatid segregation (gene ontology-derived, C5 gene sets) (Figs 5E and G, and EV3E). These findings indicate that elevated concentrations of ketone bodies not only induce cardiac EC proliferation in vitro, but might also do so in an in vivo setting.
Cardiac EC proliferation in mice is transiently increased by ketogenic dietGene profiling revealed that administration of ketogenic diet alters the transcriptional landscape of cardiac ECs in a way to promote cell division. To test this hypothesis, we determined the amount of Ki-67-positive cardiac ECs in mice fed with control or ketogenic diet for 2 weeks. ECs were stained against erythroblast transformation-specific-related gene (ERG), a highly specific EC marker localized in the cell nucleus (Nikolova-Krstevski et al, 2009). This allowed co-detection of nuclear Ki-67, a classical proliferation marker. The analysis revealed that there was a significant increase in the abundance of Ki-67 positive ECs in hearts of mice kept on a ketogenic diet for 2 weeks when compared to animals fed with a control diet (Fig 6A). Incorporation of 5-ethynyl-2ʹ-deoxyuridine (EdU) into newly synthesized DNA is an alternative method to detect proliferating cells. EdU was administered orally starting from the day of switching the diet until the end of the study. This assay showed higher numbers of EdU-positive cardiac ECs in mice fed a ketogenic diet when compared to animals that had received a control diet (Fig 6B). No differences in the abundance of apoptotic cells were determined by detection of cleaved caspase 3-positive (Fig EV4A).

Figure 6. Ketogenic diet induces proliferation of cardiac endothelial cells in mice
A, B. Representative images of heart sections of animals kept on a ketogenic diet or control diet for 2 weeks stained against ERG and Ki67 or EdU. Double-positive cells are indicated by arrowheads. Quantification is of double-positive cells in heart sections per high power field (HPF). C, D. Quantification of Ki67+/ERG+ cells in heart sections of mice kept on a ketogenic diet or control diet for 4 or 6 weeks. E. Quantification of CD31+ area in heart sections of mice kept on a ketogenic diet or control diet for 4 months. F. Quantification of Ki67+ endothelial cells in several organs of mice kept on a ketogenic diet or control diet for 4 weeks (gastrocnemius and soleus muscle) or 2 weeks (brain, lungs, subcutaneous adipose tissue, liver). Scale bar: 50 μm. Data are presented as mean ± SD. n ≥ 3; *P < 0.05; **P < 0.01; ***P < 0.001 unpaired Student’s t-test. Click here to expand this figure.
Figure EV4. Endothelial cell apoptosis in hearts of mice receiving a ketogenic diet
A, B. Quantification of cleaved caspase 3+/CD31+ double-positive cells per high power field (HPF) in heart sections of C57Bl/6J mice kept on a control diet or a ketogenic for (A) 2 weeks or (B) 4 weeks. C. Representative images of heart sections of animals kept on the control or ketogenic diet for 4 weeks stained against CD31. D–G. Quantification of CD31-positive area per high power field (HPF) in heart sections of mice kept on control diet (ctrl diet) or ketogenic diet (keto diet) for 2 weeks / 4 weeks / 6 weeks / 4 months. Scale bar: 50 µm. H. Quantification of Ki67+/CD31+ double-positive cells per high power field (HPF) in B16F10 tumor sections of C57Bl/6J mice kept on a control diet or a ketogenic 10 days after tumor inoculation. I. Quantification of CD31+ vessels per high power field (HPF) in B16F10 tumor sections of C57Bl/6J mice kept on a control diet or a ketogenic 10 days after tumor inoculation. J. Tumor volume of C57Bl/6J mice kept on a control diet or ketogenic diet. Data are presented as mean ± SD. n ≥ 3; statistical significance determined using unpaired Student’s t-test.Next, we tested whether ketogenic diet would increase the number of proliferating cardiac ECs when given over a longer period. First, the number of Ki-67-positive ECs was determined after 4 weeks of feeding the respective diets. Again, there was a pronounced increase in Ki-67-positive ECs but no increase in apoptotic ECs in mice receiving a ketogenic diet (Figs 6C and EV4B). However, in mice that received ketogenic diet for 6 weeks, we could not detect in the number of proliferating ECs between the two groups anymore (Fig 6D). This indicates that ketogenic diet leads to a transient increase in cardiac EC proliferation. Consistently, we could not detect differences in the cardiac vessel density of animals kept on a ketogenic diet when compared to littermate animals kept on a control diet at different time points for up to 4 months (Figs 6E and EV4C–G). In addition, the ratio of EC to non-EC cardiac cells was not changed. As such, ketone bodies induce a transient increase in the basal proliferation rate of cardiac ECs which does not result in an expanded blood vessel network.
Ketogenic diet does not promote EC proliferation in other organsWe wondered whether the observed transient effect of ketogenic diet on EC proliferation in the heart would also occur in other vascular beds. Therefore, the ratio of Ki-67-positive versus negative ECs was determined. This revealed, that 2 weeks of ketogenic diet increased EC proliferation only in the heart (Fig 6A and B) but not in skeletal muscle (gastrocnemius and soleus), brain, liver, lung, or white adipose tissue (Fig 6F).
EC turnover is low in the quiescent vasculature of organs in the adult vasculature. However, in tumors, there is a reactivation of the embryo-fetal angiogenic program leading to high EC proliferation rates (Rafii et al, 2016; Augustin & Koh, 2017). To investigate whether ketone bodies would affect angiogenesis, B16 melanoma cells were implanted into the skin of C57Bl/6J mice, a model leading to rapid formation of new blood vessels. There were no effects on tumor EC proliferation or vessel density in mice fed a ketogenic diet. In addition, tumor growth rates were not different between both diet groups (Fig EV4H–J).
Proliferation of cardiac ECs is increased after pressure overload-induced cardiac hypertrophy in mice kept on a ketogenic dietThe data suggest that ketogenic diet transiently increases the proliferation capacity of ECs specifically in the heart. Therefore, we wondered whether this might be beneficial in the diseased heart. Hemodynamic overload in response to, for example, hypertension, aortic stenosis, or myocardial infarction, can be partly compensated by a hypertrophic response. Pathological cardiac hypertrophy induces maladaptive cardiac remodeling and dysfunction characterized by vascular rarefaction amongst others (Nakamura & Sadoshima, 2018).
We therefore investigated whether feeding a ketogenic diet in a prevention trial would affect vascular remodeling in an acute pressure overload model causing cardiac hypertrophy (Fig EV5A). Transverse aortic constriction (TAC) increased aortic flow rate within the stenosis in both diet groups to a similar degree (Fig EV5B) and led to thickening of the left ventricle 8 weeks after surgery in mice fed a control or ketogenic diet (Fig EV5C). Parameters for contractile cardiac function were unchanged between animals fed a control or ketogenic diet in the TAC-operated and the sham group (Fig 7A). In the control diet group, there were no differences in the percentage of proliferating cardiac ECs between the sham and TAC-operated animals (Fig 7B and C), indicating that, as expected, cardiac hypertrophy leads to vascular rarefaction. Indeed, mice on control diet showed lower blood vessel density in the hypertrophic left ventricular wall compared to sham-operated mice (Figs 7D and EV5D). However, mice fed a ketogenic diet had higher rates of EC proliferation in the hypertrophic left ventricle upon TAC compared to sham mice also receiving ketogenic diet (Fig 7B and C). Consistently, blood vessel density was maintained between TAC and sham animals in the group fed with ketogenic diet whereas blood vessel density declined in the group fed with control diet (Figs 7D and EV5D).
Click here to expand this figure.
Figure EV5. Schematic overview of experimental animal groups for transverse aortic constriction study and aortic flow rates
Male and female C57Bl/6J mice were randomly assigned to a diet group at 8 weeks old. After 4 weeks on the respective diet, mice in each group underwent either transverse aortic constriction (TAC) or sham surgery. Mice were afterward kept on the respective diet for eight more weeks. Quantification of aortic blood flow rates within the stenosis of C57Bl/6J mice after TAC or sham surgery. Data are presented as mean n ≥ 2. Trichrome staining of heart sections from sham and TAC-operated animals kept either on a control diet or ketogenic diet 8 weeks after surgery. Scale bar: 1 mm. Representative images of heart vasculature (CD31+ vessels) of mice kept on a control or ketogenic diet 8 weeks after TAC surgery. Scale bar: 50 µm. Data are presented as
留言 (0)