
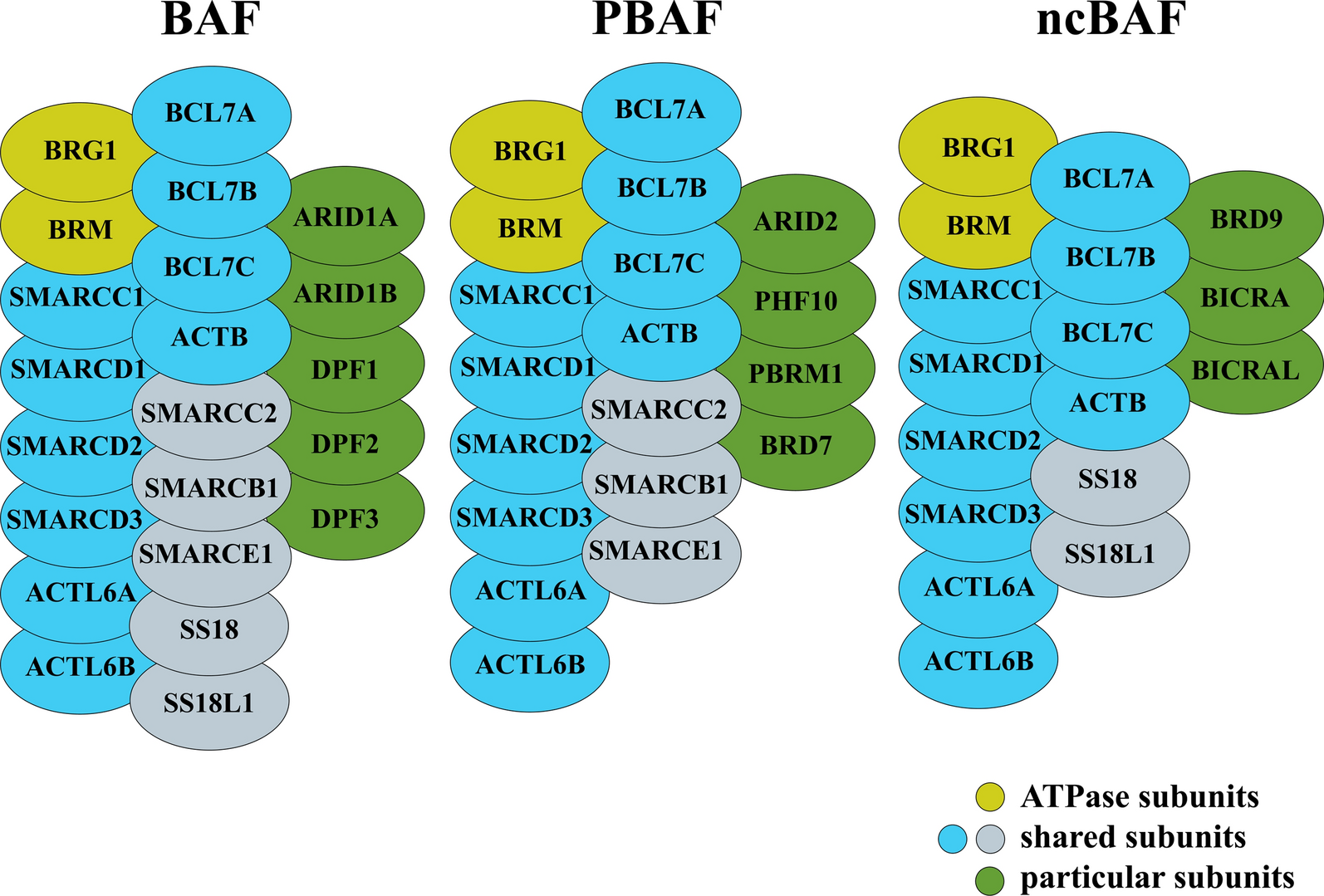
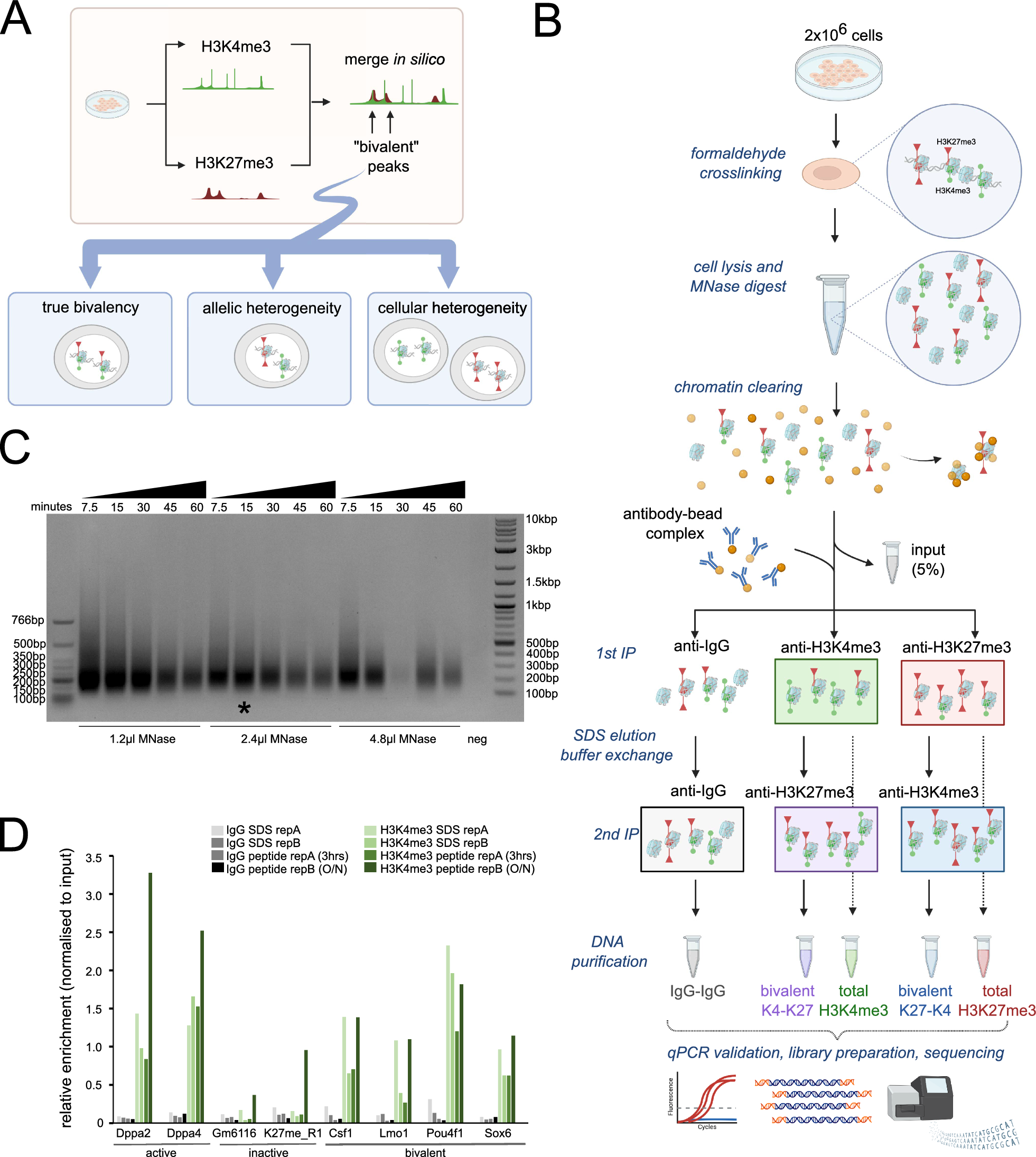
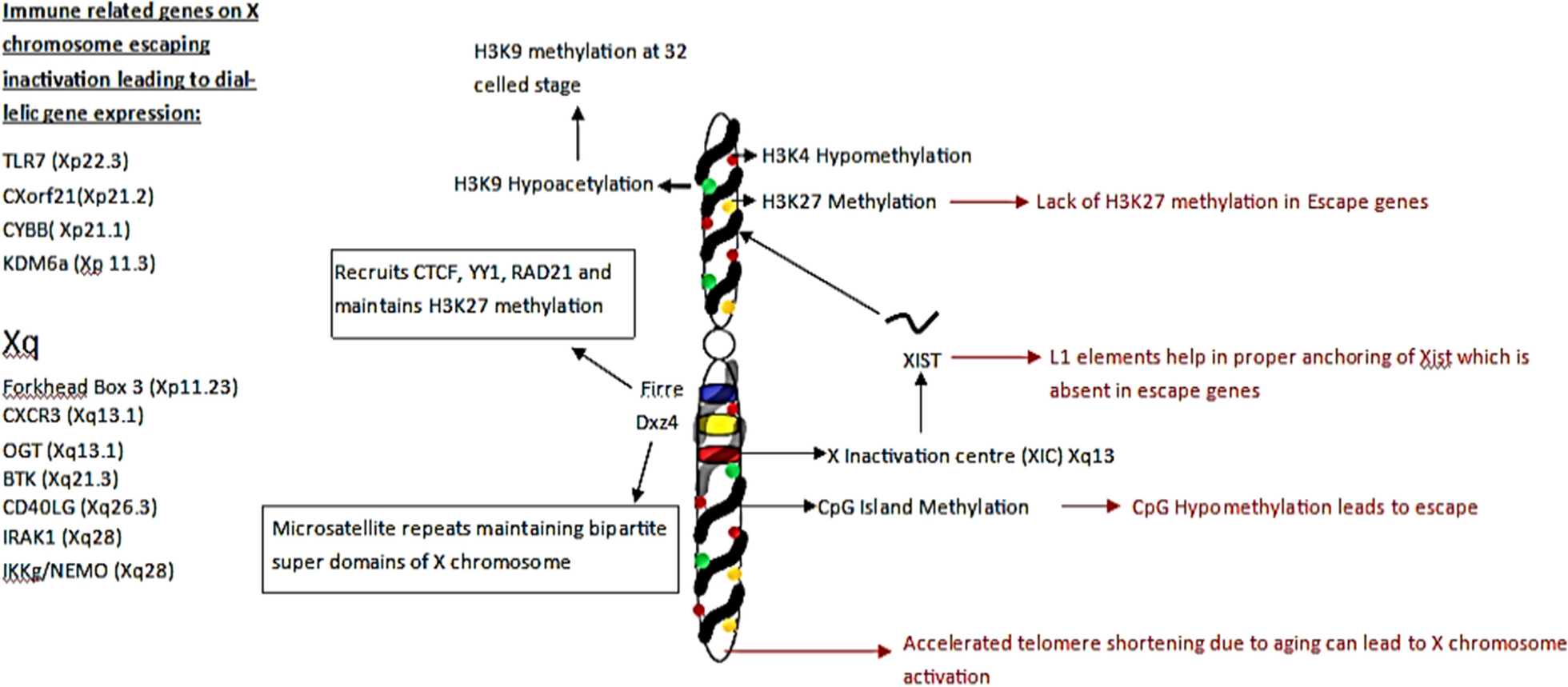
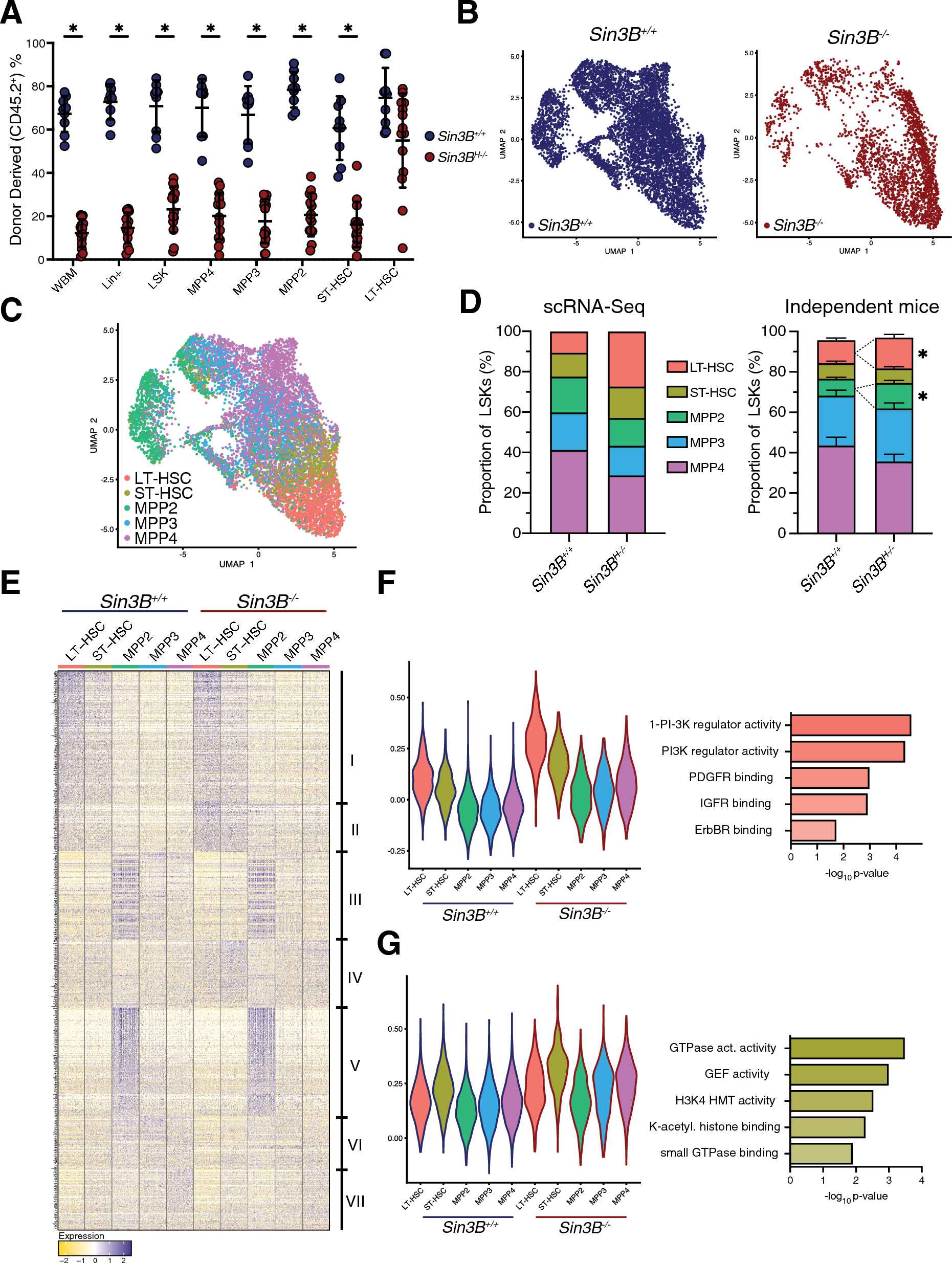
記住我
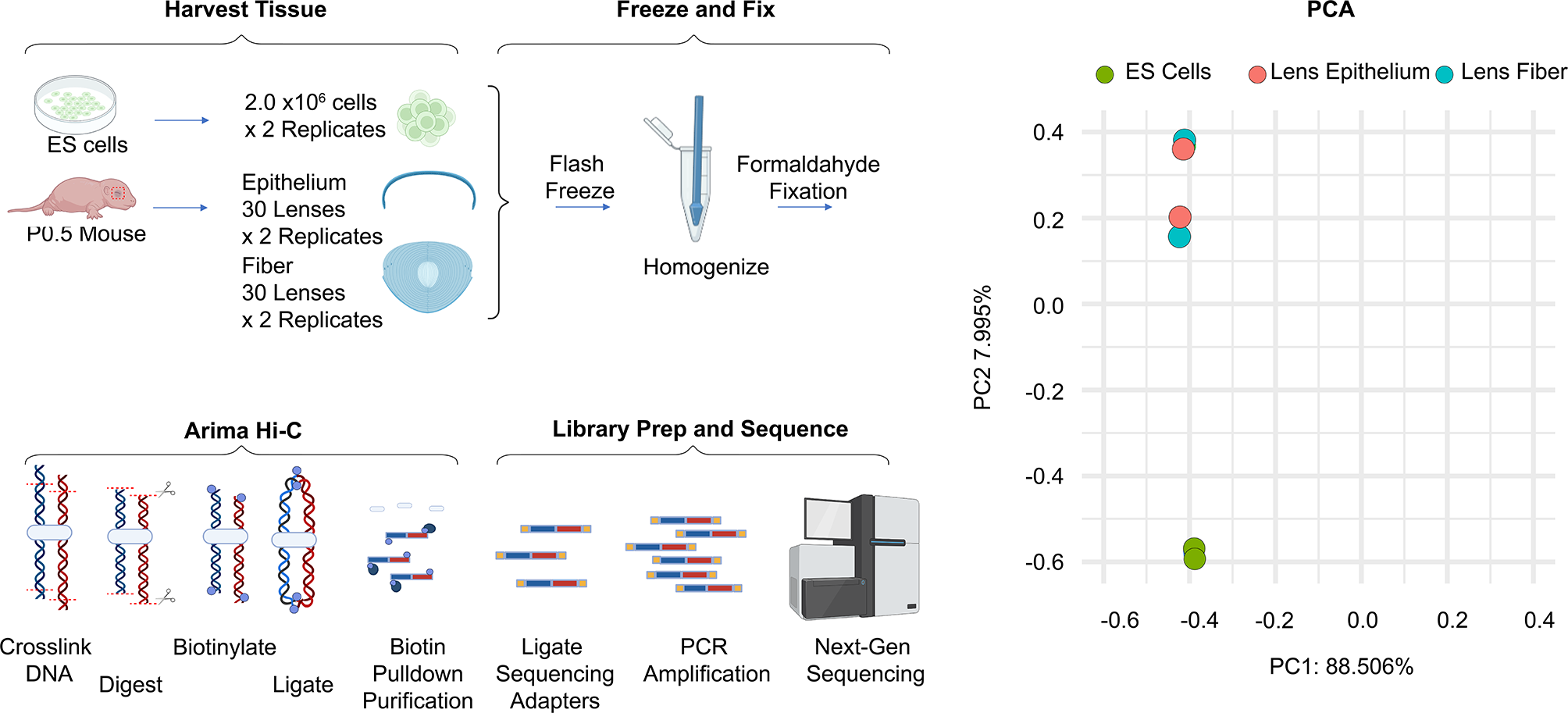


The mechanisms by which XIST functions as a cis-acting repressor of gene expression are not yet understood, particularly in humans. To follow the dynamics of XIST-mediated transcriptional repression, we examined allelic silencing upon XIST induction. While it has been shown that induction of the region of XIST encompassing Repeat A is sufficient to silence an adjacent reporter gene, it is not known what elements are necessary for XIST to silence distal genes [7, 17]. Four genes with coding SNPs megabases from the site of the XIST transgene integration into chromosome 8p were identified: CTSB (~ 5 Mb), DLC1 (~ 6 Mb), SLC25A37 (~ 17 Mb) and STC1 (~ 17 Mb). While silencing of 8p genes further away has been observed by RNA-seq analysis, these four genes were studied as they have coding SNPs and have been well-validated to consistently become silenced following XIST induction in this model [16]. Allele-specific silencing was measured by pyrosequencing of a region of the cDNA of these genes containing a SNP before and after the induction of the XIST cDNA transgene with dox. Without XIST induction the two alleles contributed approximately equally, and the strength of silencing of a specific allele was determined by its decreased representation following XIST induction. The change in allelic contribution was measured at six selected intervals between 2 and 12 days of XIST induction (Fig. 1A and Additional file 1: Table S1). Within 2 days of XIST cDNA induction, we observed a substantial reduction in allele-specific contribution across these four genes. Three of the four genes were silenced at day two to levels statistically comparable with the maximal levels observed (Fig. 1A). Only the most distal gene, STC1, differed significantly at day two of XIST induction from the subsequent levels at later time points, indicating that silencing was still underway. By the day five time point, maximal levels of allelic silencing were evident at all alleles (Fig. 1A).
Fig. 1
XIST-mediated silencing depends on Repeats A and F as well as 3’ non-repeat domain. A The change in allelic contribution over time of four 8p genes repressed by the induction of the XIST cDNA construct in the HT1080 cell line are shown. Three biological replicates were tested for each time point and significance was calculated using a student t-test relative to 5ddox. B The change in allelic contribution (for the same genes as in A) caused by 5 days of induction of XIST deletion constructs are shown normalized to the full-length XIST construct. A value of 1 indicates that the strength of silencing was comparable to Full XIST, a value of 0 indicates no silencing occurred. Deletion constructs where all 4 alleles statistically differed from Full XIST by t-test with multiple testing correction are indicated. C FISH images of XIST (green) and Cot-1 (red) and DAPI (blue) used to exemplify the formation of a transcriptionally inert “Cot-1 hole” in Full XIST HT1080 cells treated with dox for 5 days. D The depletion of Cot-1 RNA at the site of XIST RNA was calculated as the standard score (z-score) for each of the 60 cells examined at 2, 5 and 10 days of induction. Cell populations were blinded through the process and the distribution of conditions were statistically compared using a Mann–Whitney test with multiple testing correction. There was no significant change across the timepoints. E Each deletion construct induced for 5 days was tested for its relative depletion (z-score) of Cot-1 RNA hybridization at the site of XIST. Samples were blinded and statistically compared to Full XIST using a Mann–Whitney test with multiple testing correction. A–E The thresholds for statistical significance of all p values are: * < 0.05, ** < 0.01 and *** < 0.001, with these thresholds adjusted based on the number of tests
We then assessed the impact of nine different CRISPR-mediated deletions spanning the full XIST transgene (Fig. 1B and Additional file 1: Table S2 (see ref. [13]). The change in allelic ratio observed for each transgene deletion cell line was normalized to the change in allelic ratio for full-length XIST and presented as strength of silencing, where a value of 1 indicated comparable silencing to Full XIST and 0 indicated no change in allelic contribution from the pre-induction levels. The ~ 1.7-kb region spanning both Repeats A and F was found to be critical for silencing of all four alleles examined as there was no change in relative allelic contribution following induction of either the Delta A or Delta F constructs (Fig. 1C, Additional file 1: Table S3). Additionally, a ~ 630-bp region at the 3’ end (Delta 3’) also significantly impacted allele-specific silencing. Deletion constructs previously generated by FLP-mediated re-integration were consistent with the CRISPR deletion constructs with loss of the large 3.8 kb internal region of exon 1 not affecting silencing but the loss of the region corresponding to the post exon 1 sequence (3.6 kb) replicating the deficits in silencing observed in the Delta 3’ construct (Additional file 2: Fig. S1).
The ability of XIST to form a Cot-1 hole (a transcriptionally inert three-dimensional domain within the nucleus) was examined by RNA FISH using Cot-1 as a probe (Fig. 1C) which hybridizes to the heteronuclear transcripts of the genome [19]. Quantification was performed by line measurement of fluorescence intensity across nuclei and calculating the standard score (z-score) of Cot-1 fluorescent intensity at the XIST RNA cloud, as previously described [13]. The transcriptionally inert domain generated by XIST had not reached its maximal level by day 2 (median z-score = 1.06, Fig. 1D). This suggested that the nuclear reorganization mediated by XIST to create a transcriptionally inert domain lagged slightly behind the allele-specific silencing observed in Fig. 1A. Across the various transgene deletions, again the largest impact was from regions encompassing Repeats A and F (p < 1.0 × 10–13, Fig. 1F and Additional file 3: Fig. S2). Surprisingly, we observed that the ~ 1-kb region encompassing Repeat F (z-score = 0.182) to be slightly more important than Repeat A (z-score − 0.483) for forming the transcriptionally inert domain (p = 2.1 × 10–05). No other region of XIST was found to be critical for the formation of this transcriptionally inert nuclear domain suggesting that the 5’ region of XIST is specifically essential for this process.
The two methods of measuring transcriptional repression confirmed the critical role for the A repeat region in silencing, and also demonstrated a similarly critical role for the nearby F repeat region. However, there were also surprising differences between the approaches. The dispensability of the 3’ most region of XIST for the formation of a transcriptionally inert domain suggested that additional factors at the 3’ end of XIST are contributing specifically to the silencing of actively expressed genes. Genic silencing was accomplished quickly after XIST induction, while Cot-1 suppression accumulated over time. We have previously seen that recruitment of PRC1/2 and their affiliated marks is maximal at day 5 or later [13], so the recruitment of other heterochromatic marks and the loss of euchromatin may contribute to Cot-1 suppression.
Acetylation most resembles silencing in requirement for critical XIST domainsSince silencing occurred before maximal recruitment of the heterochromatic marks (H3K27me3 and ubH2A) previously examined [13], we explored additional chromatin marks or proteins described to be established by the XIST RNA. As previous studies have linked the loss of histone acetylation with the loss of transcriptional activity, we set out to examine the dynamics of histone acetylation loss at the nuclear domain of XIST and determine whether we could observe a depletion of histone acetylation using this model system and to identify the regions of XIST critical for this deacetylation. Depletion was detectable by day 2 (z-score = − 1.62), but was greater by day 5 (z-score = − 2.16) by which point it plateaued and did not accumulate further by day 10 (z-score = − 2.17, Fig. 2A and Additional file 1: Table S1). Analysis of the transgene deletions for their depletion of histone acetylation (H3K27ac) at the XIST focal accumulation was performed using the same method used to examine the depletion of Cot-1 and it was observed that only loss of the A repeat in the Delta A deletion disrupted XIST-mediated histone deacetylation (z-score = − 0.17, p = 2.6 × 10–18, Fig. 2B, Additional file 4: Fig. S3 and Additional file 1: Table S4). Previous studies examining the regions of XIST critical for the recruitment of the PRCs [13] had observed numerous interdependent domains across the length of XIST, suggesting that HDAC recruitment and/or activation by XIST operated at least semi-independently of the PRCs.
Fig. 2
The regions of XIST critical for histone deacetylation and enrichment of H4K20me1 and CIZ1. A Example images showing typical depletion of H3K27ac (red) at the site of XIST RNA after five days of induction in the HT1080 cell line and z-score s showing the depletion of H3K27ac over time by XIST induction. B Each deletion construct induced for 5 days was tested for its relative depletion (z-score) of H3K27ac at the site of XIST (59–60 cells per construct). The distributions of z-scores for each construct are shown in boxplots with significance calculated relative to the Full XIST control. C Similar to A, example images and time point analysis were performed for the enrichment of H4K20me1 at the site of Full XIST expression in the HT1080 cell line. D Similar to B, the deletion constructs were tested for their enrichment of H4K20me1 after 5 days of XIST induction and the population distributions of the 60 cells analysed are shown in boxplots. E Example images of the enrichment of CIZ1 at the site of XIST and the population distribution of the enrichment profile of cells (z-score) at various induction timepoints. F The plotted enrichment profiles of CIZ1 at XIST in each of the deletion constructs. A–F All samples were blinded throughout the analysis process and statistical significance calculations were performed using the Mann–Whitney test with the thresholds for statistical significance of all p values being: * < 0.05, ** < 0.01 and *** < 0.001, with these thresholds adjusted based on the number of tests
In addition to the more extensively studied pathway of XIST-mediated histone deacetylation, we wished to examine the enigmatic role of H4K20me1, established by KMT5A. While one of the earliest marks observed to be enriched on the inactive X chromosome [20], it has been studied less than the other marks described so far and little is known of the pathway regulating its activity. Analysis of H4K20me1 across three time points revealed that the mark plateaued statistically at day five as reported for the PRC mediated marks described previously (5- and 10-day z-score s = ~ 2.5, Fig. 2C and Additional file 5: Fig. S4 and Additional file 1: Table S1). The regions of XIST crucial for H4K20me1 enrichment were examined using the XIST transgene deletion cell lines. The enrichment profiles for the 60 cells examined in each case were compared to the Full XIST control population (Fig. 2D and Additional file 1: Table S4). We observed that loss of the non-repeat region between the C and D repeat region disrupted the enrichment of H4K20me1 by XIST, as did the loss of the region including Repeat E. Loss of any other region of XIST did not impair enrichment of H4K20me1 (mean z-score s ≥ 2, Fig. 2D).
Repeat E has been reported to be important for XIST localization through recruitment of CIZ1 and associated condensate-forming RNA binding proteins [21]. We thus extended our analysis to include CIZ1 and observed a strong co-localization with XIST that remained constant across all time points (Fig. 2E and Additional file 1: Table S1). Consistent with other studies, CIZ1 recruitment was solely dependent on the presence of the E repeat region (z-score = − 0.03, p = 3.6 × 10–21 Fig. 2F, Additional file 6: Fig. S5 and Table S4). CIZ1 requires only this single domain of XIST to be recruited and was fully enriched by the earliest time point (2-day z-score = 12.9, Fig. 2E and F, Additional file 1: Table S1 and S4).
Overall, examination of the domains of XIST required to recruit features showed that silencing and loss of acetylation both required repeat A; while H4K20me1 and CIZ1 recruitment to the XIST domain had no overlap with domains required for silencing, although they both required the E repeat region. Enrichment of the majority of proteins and histone marks to the XIST domain did not appear to be critical for silencing. In addition to the well-studied Repeat A region, we also observed that the ~ 1-kb region encompassing Repeat F and a 3′ 630-bp region were critical for silencing. While those domains were not required for H4K20me1 or CIZ1 recruitment, we previously demonstrated that H3K27me3 enrichment required the Repeat F surrounding region, while the 3’ region was critical for ubH2A recruitment [13].
Chromatin remodelling by PRC2 and KMT5A impacts XIST-mediated silencingAs previous studies have suggested linkages between the ability of XIST to induce gene silencing and chromatin-remodelling pathways, we set out to examine what connections exist within this system in which XIST is induced in differentiated cells. The domains of XIST required for silencing overlap domains required for the chromatin features established by XIST, however it remains to be determined whether these pathways are interdependent. To test the role of the chromatin-modifying complexes previously associated with XIST-mediated activity, we used small molecule inhibitors that impacted the enzymatic activities of these complexes without disrupting the levels of the component proteins.
Cells were treated for 5 days in dox in combination with inhibitors. We anticipated that silencing might be sensitive to histone deacetylation given that acetylation required the A repeat region, which is essential for silencing in both human and mouse. Inhibitors were initially tested across multiple concentrations to determine the highest functional concentration in which the cells could survive 5 days. The H3K27ac antibodies used for IF proved unsuitable for examining levels of histone acetylation by western blotting, so we instead used H4K8ac antibodies validated and gifted to us by the Howe lab at UBC. For the broad-spectrum HDAC inhibitor TSA the highest concentration (60 nM) resulted in over a fourfold increase in histone acetylation (H4K8ac) when treated cells were compared to control populations through western blotting (Additional file 7: Fig. S6A). A dose-dependent effect of TSA was observed across the lower doses, with the 40 nM TSA treatment showing a greater than twofold relative increase in histone acetylation compared to controls (Additional file 7: Fig. S6A). Despite the increase in histone acetylation detected by western, analysis of allelic silencing mediated by 5 days of full-length XIST expression from 8p demonstrated no perturbation in any of the genes examined and did not significantly disrupt XIST expression (Fig. 3A and Additional file 8: Fig. S7 and Additional file 1: Table S5). In mouse development it has been suggested that XIST-mediated silencing via the A domain depends upon HDAC3, and thus we used RGFP966, previously shown to specifically inhibit HDAC3. Again, no reduction in silencing was observed at concentrations in which cells were viable for 5 days though a subtle but statistically significant increase in the strength of silencing of CTSB, SLC25A37 and STC1 was observed (p < 4.9 × 10–5, Fig. 3A and Additional file 1: Table S5). Treating cells with HDAC inhibitors in the absence of dox did not affect the allelic ratios (Additional file 9: Fig. S8).
Fig. 3
The role of chromatin modification on XIST-induced gene silencing. A The ability of Full XIST to induce allele-specific repression at four distal genes is shown when cells were treated with chemical inhibitors for XIST-associated chromatin remodelers. Three biological replicates were tested for each condition shown and statistical significance of a treatment’s effect on XIST-induced silencing was calculated using a t-test with multiple testing correction. The concentration of chemical inhibitors in media along with the factor inhibited are listed, and the key for the genes assessed is beside panel B. B Test of whether HDACs contribute to silencing if used at much higher concentrations for only 2 days, as longer exposure was lethal to cells. Strength of silencing for these higher HDACi treatments was normalized and compared to uninhibited 2ddox Full XIST by t-test. C Example images depicting XIST (green) mediated formation of a transcriptionally inert domain (Cot-1, red) over the course of five days of induction combined with chemical inhibition. D Boxplot showing distribution of Cot-1 depletion (z-score s) across populations of 60 cells, following 5 days of chemical inhibition and Full XIST induction. All conditions were blinded through the process and statistical comparisons of population distributions of z-score s performed using the Mann–Whitney test. A–D Chemical inhibition was performed using the following: TSA (HDACi), RGPF966 (HDAC3i), ryuvidine (KMT5A), GSK343 (PRC2i) and PRT4165 (PRC1i). The thresholds for statistical significance were all corrected for multiple testing correction from * < 0.05, ** < 0.01, *** < 0.001 (p values listed individually in Additional file 1: Table S5)
To explore whether other chromatin modifications had any impact on silencing, we also used ryuvidine, a known KMT5A inhibitor [22], which at 2 uM reduced H4K20me1 levels dramatically to 25% of control levels while the highest dose used, 4uM, reduced levels to 3% of control levels (Additional file 7: Fig. S6B). Both doses resulted in a marked increase in XIST levels, though these failed to reach statistical significance (Additional file 8: Fig. S7). At 2 uM ryuvidine there was limited impact on silencing for three of the four genes examined (p < 2.2 × 10–4); with the closest gene (CTSB) still able to silence (strength of silencing = 86%, p > 0.33, Fig. 3A, Additional file 1: Table S5). At the higher dosage of 4 uM ryuvidine the loss of silencing was significant for all four genes, although still less for CTSB (p = 8.3 × 10–3; Additional file 1: Table S5).
We also inhibited both the polycomb complexes as previously [13] and now examined the effect on gene silencing. Our previous examination of the PRC2 inhibitor GSK343 had shown impact on XIST levels at the highest doses (Additional file 8: Fig. S7), so here we examined the 2.5uM and 5uM concentrations (Fig. 3A and Additional file 1: Table S5). At 2.5 uM, we observed a significant effect on XIST-mediated gene silencing while halving global H3K27me3 levels and a near total loss of silencing at the higher dose of 5 µM when H3K27me3 levels were ~ 25% of pre-treatment levels (p < 6.0 × 10–4, Fig. 3A, Additional file 1: Table S5, Additional file 7: Fig. S6C). Inhibition of PRC1 using the chemical inhibitor PRT4165 across 25uM and 50 µM concentrations for 5 days produced a dose-dependent decrease in levels of ubH2A to 55% of control levels at 50 µM (Additional file 7: Fig. S6D). Higher concentrations of PRT4165 resulted in extensive cell death. No substantial effect of this inhibitor at either concentration was observed either on the ability of XIST to induce allelic silencing or in levels of XIST as described previously (also, Additional file 8: Fig. S7A). The weak decrease in strength of silencing of STC1 was significant (p < 2.8 × 10–4, Additional file 1: Table S5).
Previous studies have associated the ability of XIST to remove the acetylation marks that promote active transcription with the silencing induced by XIST. As we had observed HDAC inhibition at five days of treatment did not weaken XIST-mediated silencing, we examined whether higher doses at the 2-day time point could yield insights into whether XIST-mediated silencing was dependent on deacetylation. None of the treatments resulted in a statistically significant effect on XIST levels (Additional file 8: Fig. S7B). Neither the 5uM or 10uM treatment of the HDAC3-specific inhibitor, RGFP966, impacted allelic silencing statistically once multiple testing correction was introduced (p-values > adjusted threshold of 8.3 × 10–3), further suggesting that HDAC3 catalytic activity is not essential for silencing in this context (Fig. 3B, Additional file 1: Table S5). Treatment with higher concentrations of the broad-spectrum HDAC inhibitor, TSA, caused a dosage and distance-dependent effect on XIST mediated silencing (Fig. 3B, Additional file 1: Table S5). The two most distal genes, SLC25A37 and STC1, both differed significantly in their allelic distribution at all doses of TSA examined compared to controls (p < 8.9 × 10–6, Additional file 1: Table S5). At the two highest treatment doses, 200 nM and 300 nM, silencing at the next most distant gene, DLC1, was significantly attenuated (p < 3.6 × 10–4, Additional file 1: Table S5). Silencing of the closest gene to the XIST construct, CTSB, remained statistically comparable across all treatments, indicating that loss of HDAC activity did not impact the ability of XIST to repress this gene.
The ability to form a transcriptionally depleted compartment in the space occupied by XIST RNA (visualized with RNA specific fluorescent probes) was monitored with RNA FISH using Cot-1 RNA as a probe. This formation of the Cot-1 hole was also impacted by loss of the A repeat region of XIST, but more strongly impacted by loss of the region surrounding the small F repeat, suggesting some independence from the genic silencing monitored by pyrosequencing. We thus monitored the impact of these chemical inhibitors on formation of the Cot-1 hole at day 5 of XIST induction and observed it to be remarkably robust to all treatments, with no treatment showing a statistically significant weakening of depletion (Fig. 3C and D, Additional file 1: Table S6). Overall, we observed that the establishment of a Cot-1 hole was not particularly sensitive to disruption of individual chromatin marks, while spread of genic silencing up to 17 Mb was particularly sensitive to reduction in PRC2 enzymatic activity. Distance-dependent effects were observed for KMT5A and HDAC inhibition. In contrast to recent literature in mouse cells, in these human somatic cells, the impact of HDAC inhibition was not specific to HDAC3 [10, 23]. We have previously identified dependencies of MacroH2A recruitment on PRC2 activity and SMCHD1 recruitment on PRC1, implicating considerable interactions between pathways establishing XIST-dependent heterochromatin.
Interdependencies between XIST-induced chromatin-remodelling pathwaysIn order to refine potential pathways through which XIST modifies the chromatin and epigenetic profile of its chromosome of origin, we set out to identify how the various chromatin-modifying complexes affect one another. Maximal levels of enrichment for most heterochromatin features required 5 days of XIST induction as described previously (Fig. 2A–C, [13]), with only MacroH2A still accumulating at day 5. To examine what role the catalytic activity of the various chromatin-modifying factors analysed so far had on overall chromatin architecture, we incubated cells with a combination of doxycycline and chemical inhibitors as described and performed IF-FISH analysis on populations of 60 cells. Inhibition of HDAC3 in the HT1080 Full XIST cells with 1uM of RGFP966 surprisingly showed no significant impact on the z-score of depletion of H3K27ac under the XIST signal (z-score = − 1.87, p = 0.42, Fig. 4A), despite raising the genome average levels of acetylation as monitored by western blotting ~ 2.9-fold (with H4K8ac; Additional file 1: Table S7). This suggests that the balance of depletion at XIST keeps pace with gains in acetylation. Inhibition of HDAC3, however, dramatically affected the enrichment of H3K27me3 (z-score = 0.63, p = 1.7 × 10–11), simultaneously reducing H3K27me3 genome-wide by 50% as seen by western (Additional file 7: Fig. S6C). Further, this treatment completely prevented MacroH2A enrichment (z-score = − 0.11, p = 6.1 × 10–18, Fig. 4A), which we had previously shown to be dependent on H3K27me3. Inhibition of HDAC3 resulted in a subtle rise in the average enrichment of H4K20me1 at the site of XIST (z-score = 3.44, p = 8.19 × 10–4) indicating that HDAC3 activity clearly is not essential for H4K20me1 enrichment. Inhibition of HDAC3 had no discernable negative effect on the enrichment of ubH2A and SMCHD1 (Fig. 4A).
Fig. 4
Examining the connections between XIST-dependent chromatin modifiers. A–D The relative distribution of chromatin features following chemical inhibition (59–61 cells each) are shown plotted according to the chemical inhibitor used. The median z-score of uninhibited Full XIST cells is shown as a dotted red line to provide a point of comparison. All conditions were blinded throughout the process and comparisons were made using the Mann–Whitney test with multiple testing correction (p-values * < 0.05, ** < 0.01, *** < 0.001). A Cells were treated with 1uM RGFP966 to inhibit HDAC3. B Cells were treated with 60 nM TSA to broadly inhibit HDACs. C Cells were treated with 4uM of ryuvidine to inhibit KMT5A. D Cells were treated with either PRC2 inhibitor GSK343 (5 µM) or PRC1 inhibitor PRT4165 (50 µM). E Summary of the functions of XIST domains with connections between the various chromatin-remodelling factors studied (created with BioRender.com). The hashed colours depict the dependence of MacroH2A on PRC2 activity and SMCHD1 on PRC1. Pointed arrows indicate activation or contribution to a process, flat-headed arrows indicate inhibition. Solid arrows indicate an essential role while dotted arrows indicate effects that are only modestly contributing to a process. The depiction on the right shows our model of how the A, F and E repeats contribute to gene silencing and the spread of H3K27me3. The Repeat F region is essential for aggregation to repressed domains, including pre-existing H3K27me3 regions. This brings Repeat A into proximity of expressed genes (resulting in their silencing); and brings Repeat E (and bound proteins like CIZ1) together to facilitate spread of H3K27me3 along the chromosome. However, Repeat E is not necessary for silencing, and Repeat A is not necessary for H3K27me3 recruitment. Repeat E facilitates the activation and/or recruitment of PRC2 to promote the spread of H3K27me3
We next set out to compare what effect broad-spectrum inhibition of HDAC activity would have on XIST activity using the chemical inhibitor TSA. We tested the ability of XIST to remodel its chromatin in the highest tolerable concentration of TSA (60 nM) over 5 days while XIST was being induced with doxycycline. As observed with HDAC3-specific inhibition, broad-spectrum HDAC inhibition had no effect on the relative depletion of H3K27ac at the XIST RNA cloud (z-score = − 2.0, p = 0.199) despite a roughly fourfold increase in global histone acetylation (Fig. 4B, Additional file 7: Fig. S6A). Broad spectrum inhibition of HDAC activity did not discernibly effect either H4K20me1 or H3K27me3 enrichment, however we did observe a decrease in the magnitude of MacroH2A enrichment (z-score = 1.3, p = 3.51 × 10–4) and ubH2A (z-score = 2.4, p = 2.66 × 10–3) compared to the uninhibited control treatments (Fig. 4B and Additional file 1: Table S7). Interestingly, the drop in MacroH2A observed by both HDAC3i and broad HDACi was not observed with a similar decrease in H3K27me3 directly caused by 2.5uM of the PRC2-specific inhibitor GSK343 (Fig. 4A and B, [13]). A significant and unexpected increase in SMCHD1 enrichment was observed at the XIST RNA cloud (z-score = 5.1, p = 2.53 × 10–5, Fig. 4B). While beyond the scope of the current study, it is possible that the increased levels of global histone acetylation caused by broad HDAC inhibition resulted in a disproportionate decrease in SMCHD1 occupancy across the nucleus compared to the XIST loci.
As inhibition of KMT5A catalytic activity had a distance-dependent effect on silencing (Fig. 3A), yet no overlap with the domains needed for silencing, we examined the role of KMT5A on chromatin remodelling using the 4uM dose of ryuvidine found to dramatically reduce global H4K20me1 levels (Additional file 7: Fig. S6B). As anticipated, KMT5A inhibition effectively prevented any signs of H4K20me1 enrichment at the XIST RNA cloud (z-score = 0.26, p = 3.2 × 10–14, Fig. 4C) indicating that KMT5A activity was essential for the enrichment of H4K20me1. An effect of this inhibition was observed in the weakening of H3K27ac depletion at the site of XIST (z-score = − 1.6, p = 3.2 × 10–4, Fig. 4C, Additional file 1: Table S7). Inhibition of H4K20me1 produced a statistically significant weakening of the enrichment of ubH2A but did not completely abolish enrichment (z-score = 1.68, p = 1.5 × 10–8, Fig. 4C). Taken together these results suggest that KMT5A-deposited H4K20me1 operates through mechanisms primarily independent of the other factors examined here.
Previous studies examining the role of PRC2 and PRC1 on XIST activity in this model system found that these two complexes operate independently, but are each critical for the recruitment of subsequent heterochromatin factors MacroH2A and SMCHD1, respectively [13]. We set out to identify whether the inhibition of either of these polycomb complexes affected the enrichment of H4K20me1 or the deacetylation of chromatin at the XIST RNA cloud. We performed IF-FISH for H4K20me1 and H3K27ac, respectively, in treatments with either 5uM of PRC2 inhibitor GSK343 or 50 µM of PRC1 inhibitor. Neither treatment weakened the enrichment of H4K20me1 or weakened the depletion of H3K27ac at the XIST RNA cloud (Fig. 4D and Additional file 1: Table S7). Inhibition of PRC1 resulted in a moderate increase in the enrichment of H4K20me1 (z-score = 3.07, p = 3.1 × 10–4). These results suggest that the role of polycomb complexes is dispensable for histone deacetylation and H4K20 monomethylation. Enrichment of CIZ1 at the XIST RNA was also observed to be recruited independently of any of the chromatin modifiers examined here, supporting the idea that its association is not dependent upon chromatin remodelling (Additional file 10: Fig. S9).
留言 (0)