The present study explored the neural mechanism of VAS by a trial-by-trial modified visual 1-back task with nonverbal stimuli and nonverbal responses to minimize the possible influence of linguistic processing. Meanwhile, we took the modulation of target presence into account during fMRI data analyses, and compared the multielement processing to the single-element processing separately within the target-absent and target-present conditions. In the combination of confirmatory and exploratory analyses, we consistently found that visual simultaneous processing was closely related to the activations in regions belonging to VAN (i.e., bilateral IFGs) and DAN (i.e., LSPL). Especially, LSPL showed greater activation than bilateral IFGs, suggesting more involvement of DAN during VAS-related processing. Moreover, significant suppressions on bilateral TPJs were observed during multielement processing only in the target-absent condition, revealing the close relationship between TPJs and inhibition processes; meanwhile, LITG showed greater activation in target-present condition than that in target-absent condition, suggesting its possible role in general selection of attention.
4.1 A special role of left superior parietal activities in VAS-related processing
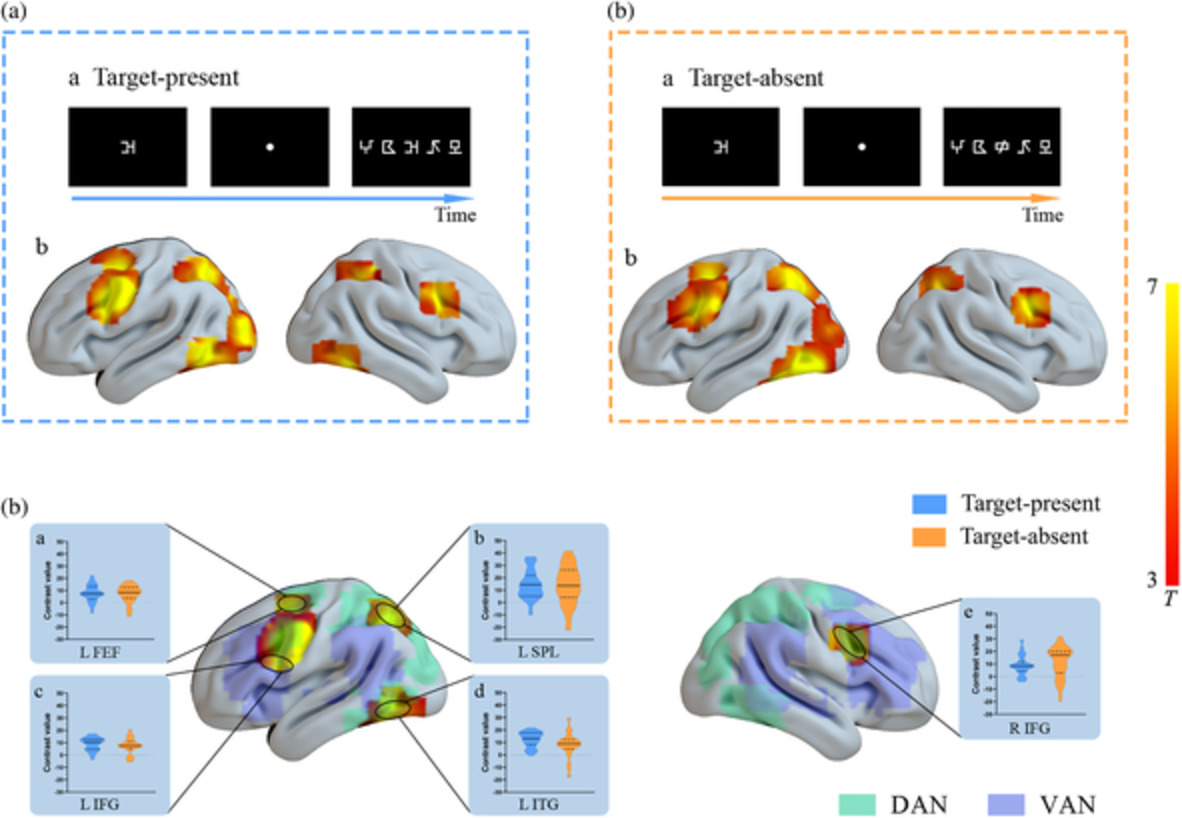
LSPL was found to be significantly activated in the target-absent and target-present conditions to similar extents, indicating that this brain region may be critical during visual simultaneous processing. Moreover, LSPL exhibited greater activation than RSPL, revealing a left lateralization in superior parietal activities which was inconsistent with previous literature reporting that VAS corresponded to brain activations in bilateral SPLs (Lobier et al., 2012, 2014; Peyrin et al., 2012; Valdois et al., 2019) with showing a right-lateralized trend (Peyrin et al., 2011; Reilhac et al., 2013; Valdois et al., 2014). These previous studies on VAS (e.g., Peyrin et al., 2011, 2012; Reilhac et al., 2013) always reported the brain activations regarding the comparisons between the multielement and the single-element conditions without considering the possible influence of the target item’s position in a string, in which overt attentional shifts might be unintentionally implicated, even though the 200-ms duration for presenting the string stimulus reduced the possibility of overt attentional shifts during the string processing (Carrasco & Hanning, 2020; Lallier, Carreiras, Tainturier, Savill, & Thierry, 2013; Talcott & Gaspelin, 2020). By contrast, we took the target presence into account during analyzing the neuroimaging data, and examined brain activities especially about the comparison between the condition when the target item appeared at the center position of a string and the single-element condition to reflect the VAS-related processing with balancing the target position, which probably relied more on covert shifts instead of overt shifts. Bilateral SPLs have been suggested to play distinct roles in spatial attentional shifts and sustained attention, and especially the responses of RSPL were related to the overt shifts but not the covert shifts whereas LSPL exhibited significant activation in covert as well as overt attentional shifts (Vandenberghe, Gitelman, Parrish, & Mesulam, 2001). Since the visual 1-back task in the present study required participants to focus on the screen center, and the data analyses controlled the possible influence of various target positions, and therefore the covert but not overt attention shift might be emphasized in the present study, with inducing the greater activation in LSPL while weakening the involvement of RSPL during the visual simultaneous processing.
In addition, although bilateral SPLs belonging to DAN have been found to be involved in visual spatial analysis (Cao et al., 2010), LSPL is more greatly associated with the visuospatial processing of the characters (Deng, Booth, Chou, Ding, & Peng, 2008; Deng, Guo, Ding, & Peng, 2012), while RSPL play a crucial role in basic visual analysis regarding spatial or nonspatial attention (Park et al., 2016). Previous studies on native speakers of alphabetic languages learning the second language such as Chinese and Japanese revealed that the reading acquisition of logographic languages would bring about neural plasticity in SPLs' functions. In detail, the experience regarding learning logographic language strengthened the activation in LSPL (Deng et al., 2008), and weakened the activation in RSPL (Sakai, Kuwamoto, Yagi, & Matsuya, 2021). Some researchers indicated that since Chinese characters have complex visual forms, and Chinese reading in daily life would exercise on the abilities of visual-orthographic analysis, with resulting in more robust activities in LSPL (Deng et al., 2012; Kuo et al., 2004). Meanwhile, the experience of Chinese characters' processing would improve our basic spatial abilities, with saving our extraneous energy to attend to and to process the nonverbal visual stimuli (e.g., symbols), which may correspond to the decrease in right superior parietal activities (Sakai et al., 2021). Accordingly, it could be proposed that previous neuroimaging studies on VAS were all in the context of French, in which the reading experience of linear alphabetic scripts may not exert a significant influence on the brain function of SPLs. By contrast, the participants in the present study were all Chinese skilled readers, and their sufficient experience of Chinese characters would emphasize the role of LSPL. Moreover, the visual complexity of the current stimuli in VAS tasks seemed to be more simple than that of Chinese characters, with less involvement of RSPL.
Moreover, further position-based analyses revealed that only responses of LSPL exhibited significant differences among positions, especially, greater activation was observed when the target appeared in the outer position (e.g., the first or fifth position) of a string as compared to the condition that the target was presented in the middle position. Consistently, analysis of behavioral data showed an effect of fixation advantage in the attentional distribution during multielement processing, which was in line with previous findings (Huang, Lorusso, Luo, & Zhao, 2019; Zhao et al., 2019; Ziegler et al., 2010). The combination of the present behavioral and neuroimaging findings may suggest a possible role of LSPL in distributing attentional resources regarding VAS. This left parietal region is critical in the division of visual attention over large segments of the visual field (Robertson, Lamb, & Knight, 1988), and it is a region implicated in spatial perception, attention, and working memory/short-term memory (Foxe et al., 2016; Panichello & Buschman, 2021). Moreover, left intraparietal sulcus (including LSPL) has been found to be activated proportionally to demand on the top-down visual spatial attention with cue-induced orienting (Hahn, Ross, & Stein, 2006), and it has been suggested that controlled attentional processes were disrupted in patients with lesions centered in left posterior parietal cortex (Robertson et al., 1988). Accordingly, based on previous literature (Bosse et al., 2007; Bundesen, 1990; Bundesen, Vangkilde, & Petersen, 2015; Stefanac et al., 2019) and the current results of the behavioral and neural responses regarding the position effect, it could be proposed that during the simultaneous processing of the symbols within the window size of VAS, the distribution pattern of VAS resources was originally imbalanced with showing a middle-position advantage. Therefore, further processing the item at outer positions required more top-down attentional control as compared to that at the middle position, with evoking greater brain activities in LSPL.
4.2 Multiple roles of bilateral inferior frontal gyrus
Bilateral IFGs were significantly and consistently activated by VAS-related processing. Previous studies (Fox et al., 2006) indicated that inferior frontal cortex belonging to ventral attention network has been found to be right-lateralized. Accordingly, we further compared the inferior frontal activities between two hemispheres in each condition regarding target presence, and found greater activation in RIFG than that in LIFG in the target-absent condition, which was consistent with the right-lateralization in previous research (Fox et al., 2006); while no lateralization difference was observed in the target-present condition. According to relevant literature (Corbetta et al., 2008; Wen, Yao, Liu, & Ding, 2012), it could be proposed that when the target item was presented in a string (i.e., target-present condition), bilateral IFGs might play roles in stimulus-driven bottom-up attentional reorientation, that was, detecting all the symbols in one string instead of limiting to the attentional focus. Whereas, in the target-absent condition, no item in one string was identical with the target stimulus and a rejective response was expected to be made, meanwhile, the function of RIFG in inhibitory control may be greatly induced as compared to LIFG with reference to relevant literature (Kolodny, Mevorach, & Shalev, 2017; Silva et al., 2019). Moreover, as shown in Figures 3 and 4, bilateral IFGs in our study seemed to locate in an overlapping region between VAN and DAN, which has also been found in previous studies (Fox et al., 2006; Vossel, Geng, & Fink, 2021). This special pattern suggested the role of IFGs in shifting attention by sending bottom-up signals from VAN to DAN. From the above results, diversified roles of IFGs (especially RIFG) were observed, that was, IFGs participated not only in visual simultaneous processing but also in response inhibition.
4.3 Suppression on TPJs during multielement processing in the target-absent condition
Bilateral TPJs, ROIs belonging to VAN, were only significantly activated in the target-absent condition rather than the target-present condition, suggesting brain activations in these regions may not be associated with VAS but with inhibition, which conflicted with previous findings reporting the involvement of TPJ in VAS-related processing (Peyrin et al., 2011; Reilhac et al., 2013; Valdois et al., 2014). However, previous researches did not consider the conditions regarding the target presence, meanwhile relevant results may also be affected by linguistic processing of verbal stimuli (Peyrin et al., 2011; Reilhac et al., 2013; Valdois et al., 2014). In the target-absent condition of the present study, bilateral TPJs were less activated in multielement processing of the nonverbal stimuli as compared to the single-element processing, revealing the suppression on TPJs activities during rapid simultaneous processing, which was consistent with previous studies indicating that TPJs as “circuit breaker” interrupted ongoing processes by reorienting new stimuli (Parks & Madden, 2013) and was suppressed during detecting target from irrelevant information (Farrant & Uddin, 2015). A large number of distractor stimuli to be rejected in the multielement condition as compared to that in the single-element condition provided more possibilities for activating TPJs to reorient. Therefore, it could be inferred that to ensure the success of target detection and discrimination during VAS processing, TPJs should be greatly suppressed especially in the multielement condition. However, the lack of significant temporoparietal activities in the target-present condition may suggest that the recruitments of TPJs did not stably reflect its role in VAS processing but possibly in inhibition processes.



留言 (0)