
記住我
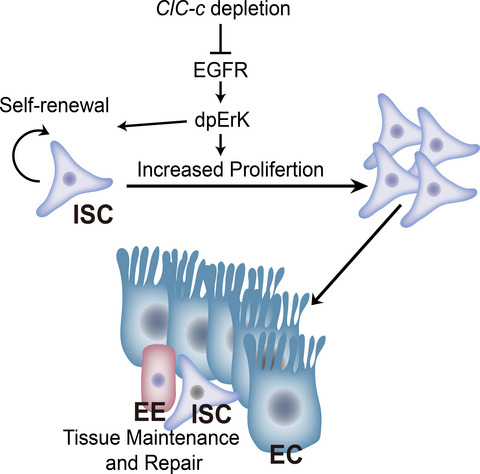



In most adult somatic organs, resident adult stem cells, whose high proliferation and differentiation abilities compensate for cell loss, are responsible for both tissue homeostasis and organ functions throughout the lifespan of the organism. Compensation of cell loss is especially important in tissues with a high turnover rate, such as the intestinal epithelium.1 Intestinal stem cells (ISCs) undergo cell division and differentiation to rapidly replenish damaged cells, thereby maintaining intestine integrity during physiological turnover or in response to damage.2, 3 Thus, characterizing the regulators and signalling pathways that control stem cell activity and maintain the critical balance between cell generation and degeneration has been one of the main focuses of developmental biology and regenerative medicine. However, how cell-fate transitions occur and how signalling pathways and transcriptional networks control these coordinated cellular changes remain largely unexplored.
The Drosophila melanogaster intestine shares many similarities with its human counterpart and thus has emerged as a powerful system to study the role of intestinal stem cells in adult tissue homeostasis and regeneration because of its relatively simple and well-characterized stem cell lineage and tractable genetic manipulation.4, 5 Drosophila ISCs are small diploid cells scattered along the basement membrane of the midgut epithelium and specifically express the transcription factor Escargot (Esg) and Notch ligand Delta (Dl).2, 3 The ISCs divide asymmetrically to generate new stem cells and either transient post-mitotic progenitor cells named enteroblasts (EBs) or enteroendocrine mother cells (EMCs). EBs progressively differentiate into polyploid enterocytes (ECs), which are responsible for the absorption of nutrients in the midgut. The other type of differentiated cells in the Drosophila intestine is enteroendocrine cells (EEs). These cells emerge from EMCs, which express markers of both ISCs (Esg) and EEs (Prospero). A high level of Notch signalling drives ISCs to produce ECs, whereas a low level drives them to produce EEs (Figure 1A).1, 4, 6-8 Under homeostatic conditions, the ISCs in Drosophila midgut are largely quiescent. The number of ISCs and progenitor cells in the midgut is relatively small and remain stable. However, these ISCs promptly undergo proliferation and differentiation when the tissue is injured.9, 10 This cellular response is essential for maintaining the epithelial homeostasis since a failure to replace lost cells may compromise tissue function and overproduction of cells may lead to cancer. Under both homeostatic and stressed conditions, ISC divisions are modulated by numerous regulators, such as Sox21a, GATAe and Hairless,11-13 and signalling pathways, such as the EGFR, Wnt, mTOR and JAK/STAT pathways,14-20 to maintain the critical balance between cell generation and degeneration. However, how these signalling pathways are regulated and integrated by which is unclear.

ClC-c is specifically expressed in stem and progenitor cells in the intestinal epithelium of the adult Drosophila. (A) Schematic diagram of the cell types and markers in the Drosophila midgut. (B) Strategy of construction of Drosophila ClC-c-3 × HA knock-in line. (C) Expression of ClC-c-3 × HA protein (red) in the esg-Gal4ts > GFP+ cells (ISCs and EBs; co-stained with GFP, green) in the R4 region of the Drosophila midgut. (D) Expression of ClC-c-3 × HA protein (red) in the ISCs (labelled by Delta staining, green) in the R4 region of the Drosophila midgut. (E and F) Expression of ClC-c-3 × HA protein (red) in the NRE-Gal4ts > GFP+ cells (EBs; co-stained with GFP, green) in the R4 region of the Drosophila midgut. Yellow arrowhead indicates the NRE-Gal4ts > GFP+ cells with small nuclei (newly formed EBs). (G) Expression of ClC-c-3 × HA protein (red) in the EEs (labelled by Prospero staining, green) in the R4 region of the Drosophila midgut. (H) Expression of ClC-c-3 × HA protein (red) in the Myo1A-Gal4ts > GFP+ cells (ECs; co-stained with GFP, green) in the R4 region of the Drosophila midgut. EB, enteroblast; EC, enterocyte; ISC, intestinal stem cell. DAPI-stained nuclei are shown in blue. Scale bar represents 10 μm
Chloride channel-3 (ClC-3) is a member of the voltage-dependent chloride channel ClC family, which functions in cell-volume maintenance, cell excitability, lysosomal acidification, ion homeostasis and Cl‾ transport across the cell membrane.21, 22 Several diseases, including cystic fibrosis, neuronal ceroid lipofuscinosis, allergic rhinitis and myocardial ischemia,23-27 have been discovered to be associated with aberrant expression of ClC-3. ClC-3 has been shown to actively participate in various cancers, facilitating the aggressiveness and metastasis of various types of malignancies, including breast, cervical, prostate, and nasopharyngeal cancers and glioma tumours.28-33 Additionally, ClC-3 is expressed in human and mouse intestinal epithelium.34 Genetic deletion of ClC-3 increases the susceptibility of mice to DSS- or TNBS-induced experimental colitis and thus preventing the intestinal recovery.34 Although ClC-3 has important roles in regulating the cell cycle, migration and apoptosis of tumour cells, its functions in intestinal stem cells are largely unknown.
This study found that ClC-c (the homologous gene of ClC-3 in Drosophila) is specifically expressed in the ISCs and EBs of Drosophila midguts. Inhibition of ClC-c expression in ISCs could inhibit their renewal. Additionally, ClC-c was found to regulate ISC proliferation through the EGFR signalling pathway. This study not only uncovers a new function of the chloride channel gene ClC-c in the modulation of ISC proliferation but also improves our understanding of ISC functions.
2 MATERIALS AND METHODS 2.1 Drosophila lines and husbandryThe following fly lines were obtained from the Bloomington Drosophila stock centre (BDSC): w1118, UAS-EGFRCA (BDSC# 9533), UAS-stg-HA (BDSC# 56562), UAS-rpr (BDSC# 5823), UAS-Rab5-GFP (BDSC# 43336), UAS-Rab7-GFP (BDSC# 42705), UAS-sSpi (BDSC# 58436), Df(3L)st-e4 (BDSC# 1317) and ClC-c RNAi (BDSC# 27034). UAS-Krn (FlyORF # F002754) was obtained from the FlyORF.
The following fly lines were obtained from the Vienna Drosophila Resource Center (VDRC): cic RNAi (v103012), cic RNAi (v103805), ClC-c RNAi (v6465) and ClC-c RNAi (v6466).
The following fly lines were kindly provided as gifts: esg-GFP, esg-Gal4, UAS-lacZ, tub-Gal4 and FTR2A lines by Dr. Allan Spradling; and esg-Gal4ts, MyolA-Gal4ts, ISC-Gal4ts and NRE-Gal4ts lines by Dr. Benjamin Ohlstein. All the Drosophila lines used in this study are listed in Table S1.
Flies were cultured in the standard medium (50 g cornmeal, 18.75 g yeast, 80 g sucrose, 20 g glucose, 5 g agar and 30 ml propionic acid per 1 L of media) at 25°C and with 65% humidity on a 12 h light/12 h dark cycle. Unless indicated otherwise, only females were used in this study.
2.2 Generation of the knockout, knock-in and transgenic fly linesTo knock out ClC-c, the following two sgRNAs located in the forepart of the CDS region of ClC-c were designed using CRISPR Optimal Target Finder with the maximum stringency (http://tools.flycrispr.molbio.wisc.edu/targetFinder/). Each sgRNA was synthesized in vitro and then subcloned into the PMD18T vector to acquire the U6 promoter. Then, the U6 promoter and sgRNA were recombined into the attB vector. This sgRNA construct was injected into the fly line y[1] w[1118]; VK00037 (BDSC# 9752) to generate ClC-c sgRNA alleles. The ClC-c sgRNA fly line was then crossed with the vas-Cas9 fly line to obtain the mutant alleles. Two independent mutant alleles were confirmed by sequencing the ClC-c locus and named ClC-cd1 and ClC-cd2.
To generate ClC-c-3×HA knock-in lines, two constructs were generated, one with two sgRNAs and the other with a homologous recombination sequence. The sgRNA construct was generated as mentioned above. To generate the homologous recombination construct, the 5ʹ homologous arm (~1 KB upstream sequence before the termination codon), 3 × HA and the 3ʹ homologous arm (~1 KB downstream sequence after the termination codon) were inserted into the PASK vector. All the final vectors were verified by sequencing and injected by Fungene Biotechnology (Beijing, China). The two sgRNAs were used to generate DNA double-strand breaks. The 5ʹ and 3ʹ homologous arms were used for homologous recombination repair. 3 × P3-RFP was used for screening. All the sgRNA sequence used are listed in Table S3.
To get UAS-ClC-c transgenic lines, the UAS-ClC-c expression vector was generated first. Full-length cDNA (RT-PCR from total mRNA of w1118 flies) of ClC-c was subcloned into the pEntry vector by using pEASY-Uni Seamless Cloning and Assembly Kit (TransGen Biotech, CU101-02) and then sub-cloned into pTW vector by using LR recombination reaction. The primers are listed in Table S3.
2.3 Clonal analysisClC-c mutant ‘Mosaic Analysis with a Repressible Cell Marker’ (MARCM) clones were generated from the ClC-c mutant line via FLP/FRT-mediated mitotic recombination. FRT2A was recombined with ClC-cd1 to generate the FRT2A, ClC-c-null mutant line, which was then crossed with the line yw, hsFLP, tub-Gal4, UAS-GFP/FM7; tub-Gal80, FRT2A/TM6B to obtain hsFLP, tub-Gal4, UAS-GFP/+; tub-Gal80, FRT2A/ FRT2A, ClC-c flies. The flies were raised at 25°C, and 5-day-old adult flies post-eclosion were subjected to 1-h heat shock twice in a water bath at 37°C. Afterwards, they were kept at 25°C and then dissected and observed at the indicated days after clonal induction (ACI).
2.4 RNA-sequencing (RNA-seq) and data analysisThe detailed process was described previously.35 The Drosophila adult midguts (R1–R5) were first dissected in phosphate-buffered saline (PBS) on ice. They were then immediately placed in a −80°C freezer. Total RNA was isolated using the isothiocyanate-alcohol phenyl-chloroform method. Whole RNA previous sequencing was carried out on the NovaSeq 6000 platform (Illumina) by Berry Genomics Corporation. The quality control was performed using FastQC (v0.11.8).
The read length of the raw RNA-seq data was 150 bp. All the reads were aligned to the Drosophila reference genome (Ensembl BDGP6 release-89). The aligned-read sam files were then converted to bam files and sorted using SAMtools. DESeq2 (v1.22.2) was used to determine the gene expression profiles of samples. P-value ≤ 0.05 following Benjamini and Hochberg correction for multiple hypothesis testing was considered to indicate differential gene expression.
2.5 Cell sorting and RT-qPCROne hundred to two hundred midguts were dissected in cold diethyl pyrocarbonate (DEPC)-treated water PBS (0.1% final solution of DEPC is added to 1x PBS) and incubated with 0.1% trypsin for 1 h at 29°C, during which the sample was softly mixed every 15 min by pipetting and inverting several times. Dissociated samples were collected after centrifugating at 4°C, 400× g for 20 min and resuspended in DEPC-PBS. The suspension was filtered with 40-μm filters. The filtered cells are then sorted using a FACS Aria II sorter (BD Biosciences) and collected into 0.5 ml DEPC-PBS. GFP+ cells in the midgut of fly strains carrying esg-Gal4ts > UAS-GFP fluorescent markers were used to sort intestinal progenitor cell population, and the midguts of w1118 flies were set as fluorescence gate.
Total gut RNA was extracted from dissected midguts by using the Trizol reagent (Invitrogen). This RNA (1 μg) was used to generate cDNA via reverse transcription, and the cDNA was subjected to quantitative polymerase chain reaction (qPCR) in a QuantStudio 5 System (Thermo Fisher Scientific). The 2−ΔΔCT method was used to calculate the expression values. The relative expression was normalized to that of Rp49. All the primers used are listed in Table S3.
2.6 Immunofluorescence microscopyAdult midguts were dissected in PBS and then fixed with 4% Paraformaldehyde for 2 h, followed by washing three times (10 min each) with PBS containing 0.1% Tween-20 (PBST). The midguts were then blocked in 0.1% BSA for 30 min at room temperature, followed by washing with PBST, and then incubated overnight at 4°C with primary antibodies diluted in PBST. After washing three times with PBST, the tissues were incubated with secondary antibodies and DAPI for 2 h at room temperature, followed by the same washing steps above. The sources and dilutions of the primary and secondary antibodies used are listed in Table S2.
Leica TCS-SP8 confocal microscope was used to acquire all the immunofluorescence images. The Leica Application Suite X (LAS X), Adobe Photoshop cc2020 and Adobe Illustrator cc2020 were used to assemble the images.
2.7 Bleomycin (BLM), dextran sodium sulphate (DSS) and paraquat (PQ) treatmentA chromatography paper was cut into 4 × 6 cm strips and saturated with 25 µg/ml BLM (Aladdin, B107423), 5% (wt/vol) DSS (MP Biomedicals, CAS: 9011–18–1, 36–50 KD) or 5 mM PQ (Aladdin, M106761) dissolved in 5% (wt/vol) sucrose. After being starved in empty vials for 1 h, flies were transferred into vials with the BLM, DSS or PQ solution–saturated chromatography paper with 5% sucrose. After 24 h (for BLM or PQ) or 48 h (for DSS) of treatment, the flies were transferred to the standard fly food, with the daily renewal of the food. Sucrose-only (5%, wt/vol) was used as a control.
2.8 TUNEL assayAdult midguts were dissected in PBS and fixed with 4% paraformaldehyde for 2 h, followed by washing with PBST. Apoptosis was assessed using the Apoptag Kit (Millipore) according to the manufacturer's instructions.
2.9 EdU incorporation assayFor EdU labelling, a chromatography paper was cut into 4 × 6 cm strips and saturated with 5% sucrose and 100 µM EdU (Baseclick). After being starved in empty vials for 1 h, flies were transferred into vials with the EdU solution–saturated chromatography paper for 24 h. The guts of the flies were then dissected in PBS and fixed with 4% paraformaldehyde for 2 h. Edu incorporation was evaluated following the manufacturer's instructions. The immunostaining procedure has previously been described.
2.10 Detection of reactive oxygen species (ROS) by using dihydroethidium (DHE)Adult midguts were dissected in PBS and then incubated in 30 μM DHE (Invitrogen) for 5 min in the dark at room temperature. The guts were then washed twice with PBS, mounted and immediately examined using a confocal microscope.
2.11 Assessment of necrosis by using propidium iodideAdult midguts were dissected in PBS and then stained with 1.5 μM propidium iodide (PI; Invitrogen) at room temperature for 15 min. The guts were then fixed with 4% formaldehyde for 20 min, washed three times with PBST, rinsed twice with PBS, mounted in Vectashield with DAPI and examined using a confocal microscope.
2.12 Fluorescence intensity statisticsImmunofluorescence images were analysed via confocal microscopy, and the fluorescence intensity statistics in the region of interest (ROI) were calculated using ImageJ. The detailed process was described previously.35
Mean fluorescence intensity = Integrated density of the background region (or cell)/Area of the background region (or cell)
Relative mean fluorescence intensity = Mean fluorescence intensity of the cell−Mean fluorescence intensity of the background region
Relative fluorescence intensity = Integrated density of the cell−Mean fluorescence intensity of the background region × Area of the cell
2.13 Quantification and statistical analysisFor all the experiments, the data were processed using GraphPad Prism and presented as average ± SD. Statistical significance was determined using the unpaired two-tailed Student's t-test unless otherwise stated in the Figure legends. For all the tests, p < 0.05 was considered to indicate statistical significance.
3 RESULTS 3.1 ClC-c is specifically expressed in the stem and progenitor cells of the adult Drosophila intestinal epitheliumTo investigate the possible role of ClC-c in the regulation of intestinal stem cells, a ClC-c-3×HA knock-in fly line was generated (Figure 1B). We used the conditional temperature-sensitive driver esg-Gal4 (esg-Gal4ts) to express the dsRNA construct (BDSC # 27034) against ClC-c and observed that the HA signal was strongly reduced (Figure S1A–C). We detected ClC-c expression in R1–R5 regions of the adult Drosophila midgut (Figure S1D). Using specific cell markers to distinguish the cell-specific expression pattern of the endogenous ClC-c protein, we found that ClC-c is specifically expressed in esg-positive cells (Figure 1C). By using esg-Gal4, UAS-GFP; Gal80ts and NRE-Gal80 (ISC-Gal4ts) flies, we found that ClC-c is expressed in ISCs (Figure S1E). Additionally, co-staining the ClC-c-3 × HA midguts for the Dl protein and HA tag confirmed that ClC-c is expressed in ISCs (Figure 1D). Next, we found that ClC-c is expressed in some newly formed EBs with small nuclei (Figure 1E, yellow arrowhead), whereas no ClC-c expression was detected in mature EBs with large nuclei (Figure 1F). These results imply that the expression of ClC-c is gradually lost during the differentiation from EBs to ECs. As predicted, no ClC-c expression was detected in mature ECs (Figure 1H) or EEs (Figure 1G). Taken together, these results indicate that ClC-c is specifically expressed in the stem/progenitor cells of the Drosophila intestinal epithelium.
3.2 ClC-c is required for ISC proliferation in the Drosophila midgutsSince the expression of ClC-c is uniquely restricted to ISCs and some newly formed progenitor cells, we first depleted ClC-c in ISCs and EBs via esg-Gal4ts driven RNA interference (RNAi) lines ClC-c RNAi#1 (BDSC# 27034), ClC-c RNAi#2 (VDRC v6465) and ClC-c RNAi#3 (VDRC v6466). The ClC-c mRNA levels in these three RNAi lines were measured via RT-qPCR (Figure S2A). Knocking down ClC-c in the esg+ cells for 7 days at 29°C significantly decreased the number of esg+ cells (Figure 2A–C, Figure S2C–E). To further determine in which cell types ClC-c exerts its functions, we knocked down ClC-c in either ISCs or EBs by using the following cell-specific Gal4 drivers: ISC-Gal4ts for ISC knockdown; and NRE-Gal4, Gal80ts (NRE-Gal4ts) for EB knockdown. We found that depletion of ClC-c in ISCs decreases the number of ISC+ cells (Figure 2D–F), whereas no effects on the number of EBs in depletion of ClC-c in NRE+ cells (Figure S2F–H). Additionally, depletion of ClC-c in EBs did not affect the number of esg+ cells (Figure S2I–K). We also tested the effects of overexpression of ClC-c on ISCs. Overexpression of ClC-c did not significantly promote ISC proliferation (Figure S2L-N). These results suggest that ClC-c might act cell-autonomously in ISCs to regulate ISC proliferation.

ClC-c is required for ISC proliferation in the Drosophila midgut. (A and B) Immunofluorescence images from the R5 region of the midgut of the control flies (A, esg-Gal4ts-driven UAS-lacZ) or ClC-c-depleted flies (B, esg-Gal4ts-driven ClC-c RNAi#1, BDSC# 27034). (C) Quantification of the number of the esg+ cells in the midguts of the control and ClC-c RNAi groups. ROI: 2 × 104 µm² area from the R5 region of the Drosophila midgut. Gut n ≥ 10; ROI n ≥ 30. (D and E) Immunofluorescence images from the R5 region of the midgut of the control flies (D, ISC-Gal4ts-driven UAS-lacZ) or ClC-c-depleted flies (E, ISC-Gal4ts-driven ClC-c RNAi#1, BDSC# 27034). GFP (green) identifies ISCs. (F) Quantification of the number of the GFP+ cells in (D) and (E). ROI: 2 × 104 µm² area from the R5 region of the Drosophila midgut. Gut n ≥ 10; ROI n ≥ 30. (G) Schematic summary of the ClC-c mutant alleles generated using CRISPR-Cas9. (H–O) Immunofluorescence analyses of the control (FRT2A, H–K) and ClC-cd1 mutant (L-O) MARCM clones (green) in the midguts 4, 7, 11 and 14 d after clonal induction (ACI). (P) Quantification of the cell number per MARCM clone in H-O. Each dot corresponds to one clone. Gut n ≥ 10; clone n ≥ 30. ISC, intestinal stem cell; MARCM, mosaic analysis with a repressible cell marker; ROI, region of interest. DAPI-stained nuclei are shown in blue. Scale bar represents 10 μm. Error bars represent SDs. Student's t-test, ****p < 0.0001
Next, ClC-c mutant alleles were generated using CRISPR/Cas9 to validate the ClC-c RNAi phenotypes. We designed a sgRNA targeting the first exon of ClC-c and isolated two independent lines, each carrying one of the mutant alleles ClC-cd1 and ClC-cd2. By sequencing the ClC-c locus, both lines were confirmed to be mutant as many base deletions occurred after position c.392 in exon 1, thus introducing a premature termination codon into this exon in both lines (Figure 2G). No progeny was obtained when these lines were crossed with the corresponding deficiency line Df (3L) st-e4, in which the entire ClC-c locus is deleted (Figure S2B). These results suggest that ClC-cd1 and ClC-cd2 are null or loss-of-function alleles.
To observe the phenotypes of the ClC-c mutants, we used the MARCM system in which both the control and ClC-c mutant clones were marked by GFP expression. The GFP-marked clones were analysed at the indicated time points after ACI. The control clones showed normal growth at days 4, 7, 11 and 14 (Figure 2H–K,P, and Figure S2O), whereas the growth of the ClC-c mutant clones was inhibited (Figure 2L–P). This result suggests that the ClC-c mutant clones have a restricted capacity to divide and generate progeny.
Taken together, these results strongly suggest a cell-autonomous role for ClC-c in regulating ISC proliferation.
3.3 ClC-c is required for midgut regenerationIntestinal stem cells confer a high regenerative capacity to the intestinal epithelium.36 They undergo rapid cell division and differentiation to replenish damaged cells, thereby maintaining tissue integrity and the proper number of differentiated cells under the homeostatic conditions or in response to stress.37 ClC-c has been shown to be involved in ISC proliferation during homeostasis. Therefore, we utilized the ClC-c-3×HA reporter line to visualize the expression level of ClC-c in esg+ cells in injured midguts. The abundance of ClC-c considerably increased in BLM (causes DNA breaks and genomic instability)-, DSS (a model for experimental inflammatory bowel disease)- or PQ (induces oxidative stress)-injured midgut epithelium (Figure 3A–E). Treatment of the Drosophila intestinal tract with BLM, DSS or PQ caused intestinal damage and induced ISC hyperproliferation as indicated by the increase in the numbers of esg+ and pH3+ cells (Figure 3F–I,N,O). However, the numbers of esg+ and pH3+ cells in the intestines of the flies with depleted ClC-c did not increase when these flies were subjected to these injuries (Figure 3J–O). These results suggest that ClC-c is essential for inducing ISC proliferation in response to damage to the intestinal epithelium.

ClC-c is required for midgut regeneration. (A–D) Immunofluorescence images from the R5 region of the midgut, showing the expression of ClC-c-3 × HA protein (red) upon Sucrose (A), BLM (B), DSS (C) or PQ (D) treatment. (E) Quantification of the fluorescence intensities of ClC-c-3 × HA in the midguts in (A–D). Each dot corresponds to one cell. Gut n ≥ 5; cell n ≥ 90. (F–I) Immunofluorescence images from the R5 region of the midgut of the control flies (esg-Gal4ts-driven UAS-lacZ) treated with sucrose (F), BLM (G), DSS (H) or PQ (I). (J–M) Immunofluorescence images from the R5 region of the midgut of ClC-c-depleted flies (esg-Gal4ts-driven ClC-c RNAi#1, BDSC# 27034) treated with sucrose (J), BLM (K), DSS (L) or PQ (M). (N) Quantification of the number of the GFP+ cells in the midguts in (F–M). ROI: 2 × 104 µm² area from the R5 region of the midgut. Gut n ≥ 10; ROI n ≥ 30. (O) Quantification of the number of the pH3+ cells in the midguts in F–M. BLM, Bleomycin; DAPI labels the nucleus (blue); DSS, Dextran Sodium Sulphate; PQ, Paraquat. ROI, region of interest. Scale bar represents 10 μm. Error bars represent SDs. Student's t-test, ****p < 0.0001. Gut n ≥ 20
3.4 Reducing ClC-c inhibits the proliferation of progenitor cells, but does not lead to apoptosis or necrosisKnocking down ClC-c in the esg+ progenitor cells and clones decreased the number of esg+ cells and resulted in small clones respectively. Apoptosis is a common reason for cell loss. To assess the possibility that the esg+ cells undergo apoptosis upon knocking down ClC-c, the TUNEL assay was performed to detect apoptotic signals after 7 days of ClC-c RNAi. We did not observe a significant increase in the number of apoptotic esg+ cells, compared with the number in the control flies (Figure 4A,B,H). Forced expression of rpr (an inducer of apoptosis) in esg+ cells was used as a positive control (Figure 4C,H). These results indicate that the cell loss caused by knocking down ClC-c in esg+ cells is not due to an increase in apoptosis.

Reducing ClC-c leads to a loss of progenitor cells due to decreased proliferation but not increased apoptosis or necrosis. (A–C) Immunofluorescence images from the R5 region of the midgut of the negative-control flies (A, esg-Gal4ts-driven UAS-lacZ), ClC-c-depleted flies (B, esg-Gal4ts-driven ClC-c RNAi) and positive-control flies (C, esg-Gal4ts-driven UAS-rpr) with TUNEL treatment. (D and E) Immunofluorescence images from the R5 region of the midgut of the control flies (D, esg-Gal4ts-driven UAS-lacZ) and ClC-c-depleted flies (E, esg-Gal4ts-driven ClC-c RNAi) with PI treatment. White arrows indicate esg+ cells. (F and G) Immunofluorescence images from the R5 region of the midgut of the control flies (F, esg-Gal4ts-driven UAS-lacZ) and ClC-c-depleted flies (G, esg-Gal4ts-driven ClC-c RNAi) with DHE treatment. White arrows indicate esg+ cells. (H) Quantification of the number of the GFP+ and TUNEL+ cells in the midgut in (A), (B) or (C). ROI: 2 × 104 µm² area from the R5 region of the Drosophila midgut. Gut n ≥ 10; ROI n ≥ 30. (I) Quantification of the number of the GFP+ and PI+ cells in the midgut in (D) or (E). ROI: 2 × 104 µm² area from the R5 region of the Drosophila midgut. Gut n ≥ 10; ROI n ≥ 30. (J) Quantification of the number of the GFP+ and DHE+ cells in the midgut in (F) or (G). ROI: 2 × 104 µm² area from the R5 region of the Drosophila midgut. Gut n ≥ 10; cell n ≥ 200. (K and L) Immunofluorescence images from the R5 region of the midgut of the control flies (K, esg-Gal4ts-driven UAS-lacZ) and ClC-c-depleted flies (L, esg-Gal4ts-driven ClC-c RNAi) with EdU treatment. (M) Quantification of the number of the GFP+ and EdU+ cells in the midgut in (K) or (L). ROI: 2 × 104 µm² area from the R5 region of the Drosophila midgut. Gut n ≥ 10; ROI n ≥ 30. (N–Q) Immunofluorescence analyses of the control (N), ClC-cd1-null (O), stg-overexpressing (P) and stg-overexpressing ClC-cd1-null (Q) MARCM clones (green) in the midgut 7 days after clonal induction (ACI). (R) Quantification of the cell number per MARCM clone in (N–Q). Each dot corresponds to one clone. Gut n ≥ 10; clone n ≥ 30. DHE, dihydroethidium; MARCM, mosaic analysis with a repressible cell marker; PI, propidium iodide; ROI, region of interest. DAPI-stained nuclei are shown in blue. Scale bar represents 10 μm in (A–G, K, L) and 25 μm in N–Q. Error bars represent SDs. Student's t-test, ****p < 0.0001. NS, non-significant (p > 0.05)
Another possibility of cell death is necrosis. Therefore, we investigated whether knocking down ClC-c causes necrosis in ISCs. Necrosis is characterized by early plasma-membrane rupture and accumulation of ROS, which can be assessed through PI staining and DHE staining respectively.38 We detected no increase in PI+ or DHE+ signals after ClC-c depletion, compared with the signals in the control (Figure 4D–G,I,J).
Taken together, these results suggest that knocking down ClC-c does not induce apoptosis or necrosis in ISCs.
During homeostasis, the stability of a stem-cell pool requires continuous self-renewal of the stem cells. Inhibition of their proliferation results in the depletion of the stem-cell pool and loss of stem cells. To determine whether ClC-c is required for the proliferation of ISCs, we performed EdU incorporation assay (a method to detect cell cycle) in esg-gal4ts > GFP flies after ClC-c RNAi and quantified the number of EdU-positive cells per midgut. After 7 d of RNAi, the number of EdU+ cells was lower in these flies than in the controls (Figure 4K–M). Furthermore, forced expression of the cell cycle regulator stg rescued the cell loss phenotype induced by ClC-c depletion (Figure 4N–R and Figure S3A–E). These results suggest that ClC-c is required in maintaining ISC proliferation and the depletion of ClC-c suppressed ISC proliferation by inhibiting the cell cycle.
The differentiation of stem cell is another possible cause of the decline in the number of stem cells. We thus investigated the effect of ClC-c depletion on ISC differentiation. The experimental results showed that ClC-c mutant did not inhibit the formation of large nuclear ECs and pros+ EEs (Figure S3F,G). Moreover, statistical data showed that the proportion of EC cells in the ClC-c mutant clone was higher than that in the control group (Figure S3H).
Therefore, we believed that the decrease in the number of stem cells caused by the loss of ClC-c is due to the other fact that ISC could not maintain stemness thus undergoing differentiation.
3.5 Depletion of ClC-c downregulates the members of the EGFR signalling in the Drosophila midgutTo identify the mechanism whereby ClC-c regulates ISC proliferation, RNA-seq was performed on dissected midguts of the flies with ClC-c depleted in esg+ cells (esg-Gal4ts > ClC-c RNAi) and control flies (esg-Gal4ts > UAS-lacZ). The expression levels of esg and DI were significantly lower in the midguts with ClC-c-depleted esg+ cells than in the midguts of the control group (Figure 5B,C). This observation is consistent with the role of ClC-c in maintaining the stemness of ISCs. Additionally, the results of the RNA-seq analyses showed that a cluster of genes related to the EGFR signalling pathway (such as Egfr, Rho, Pnt, Sox21a and Ets21C) was expressed at significantly lower levels in the ClC-c-depleted midguts than in the control midguts (Figure 5A and Figure S4A,B). Among these genes, Pnt, Sox21a and Ets21C have been reported to function downstream of the EGFR signalling pathway and regulate ISC proliferation.39-41

Depletion of ClC-c downregulates genes related to the EGFR signalling in the Drosophila midgut. (A) Volcano plots of differentially expressed genes between the ClC-c-depleted Drosophila (esg-Gal4ts-driven ClC-c RNAi) and control (esg-Gal4ts-driven UAS-lacZ) midguts. (B–G) Relative mRNA levels of the genes related to the EGFR signalling in the ClC-c-depleted Drosophila (esg-Gal4ts-driven ClC-c RNAi) and control (esg-Gal4ts-driven UAS-lacZ) midguts. Error bars indicate the SD of three independent experiments. Student's t-test, *p < 0.05, **p < 0.01, ****p < 0.0001
To confirm these RNA-seq results, RT-qPCR analyses were performed. The RT-qPCR results showed similar expression patterns to those observed in the RNA-seq data (Figure 5D–G). These findings strongly suggest that ClC-c regulates ISC proliferation through the EGFR signalling pathway.
3.6 ClC-c regulates ISC proliferation through the EGFR signallingPrevious studies have shown that the EGFR signalling pathway is vital in ISC proliferation during homeostasis or stress-induced regeneration.10, 42 The expression pattern of the EGFR effector mitogen-activated protein kinase (MAPK) in the midgut was examined by staining the midgut for di-phosphorylated Erk (dpErk), the active form of MAPK. Signals of phospho-Erk were detected in the midguts of the control flies after 24 h of BLM treatment (Figure 6A,C,E), but the ClC-c-depleted group showed an unchanged level of Erk activity in response to BLM treatment (Figure 6B,D,E). Moreover, forced constitutive expression of the EGFR receptor (UAS-EgfrCA) in ISCs was sufficient to induce hyperproliferation along the whole gut epithelium (Figure 6F,H,N). The ISC proliferation defects observed in the ClC-c-depleted flies were significantly restored by overexpressing EgfrCA (Figure 6F,G,I,N). Consistently, the overexpression of EgfrCA partially rescued the clonal growth inhibition caused by knocking out ClC-c (Figure 6O–S). Moreover, activation of the EGFR signalling by knocking down cic (an inhibitor of the EGFR signalling pathway) suppressed the cell loss induced by ClC-c depletion (Figure 6F,G,J,K,N). These data indicate that ClC-c regulates ISC proliferation through the EGFR signalling pathway. Notably, we found that overexpression of the secretory EGFR ligand spi and Krn could not rescue the cell loss induced by the knockdown of ClC-c (Figure 6L–N and Figure S5A-E). These results indicate that knocking down ClC-c might inhibit the transduction of EGFR signal from extracellular to intracellular, thereby inhibiting the activation of the EGFR signalling pathway.

ClC-c regulates the proliferation of intestinal stem cells through the EGFR signalling pathway. (A–D) Immunofluorescence images of the R5 region of the midgut of the control flies (A, esg-Gal4ts-driven UAS-lacZ) with sucrose treatment, ClC-c-depleted flies (B, esg-Gal4ts-driven ClC-c RNAi) with sucrose treatment, control flies (C, esg-Gal4ts-driven UAS-lacZ) with BLM treatment, or ClC-c-depleted flies (D, esg-Gal4ts-driven ClC-c RNAi) with BLM treatment, stained for dpErk. (E) Quantification of the number of the dpErk + cells in A–D. ROI: 3 × 104 µm² area from the R5 region of the Drosophila midgut. Gut n ≥ 10; cell n ≥ 200. (F–M) Immunofluorescence images from the R5 region of the midgut of the control (F, esg-Gal4ts-driven UAS-lacZ), ClC-c–depleted (G, esg-Gal4ts-driven ClC-c RNAi), EgfrCA-overexpressing (H, esg-Gal4ts-driven UAS-EgfrCA), EgfrCA-overexpressing ClC-c-depleted (I, esg-Gal4ts-driven ClC-c RNAi with UAS-EgfrCA), cic-depleted (J, esg-Gal4ts-driven cic RNAi), [cic and ClC-c]-depleted (K, esg-Gal4ts-driven ClC-c RNAi with cic RNAi), spi-overexpressing (L, esg-Gal4ts-driven UAS- spi), or spi-overexpressing ClC-c-depleted (M, esg-Gal4ts-driven ClC-c RNAi with UAS- spi) flies. (N) Quantification of the number of the esg+ cells in the midgut in F–M. ROI: 2 × 104 µm² area from the R5 region of the Drosophila midgut. Gut n ≥ 10; ROI n ≥ 30. (O–R) Immunofluorescence analyses of the control (O), ClC-cd1-null (P), Egfr-overexpressing (Q), and Egfr-overexpressing ClC-c null (R) MARCM clones (green) in the midgut 7 days after clonal induction (ACI). (S) Quantification of the cell number per MARCM clone in (M–P). Each dot corresponds to one clone. Gut n ≥ 10; clone n ≥ 30. ROI, region of interest; BLM, Bleomycin; MARCM, mosaic analysis with a repressible cell marker. DAPI-stained nuclei are shown in blue. Scale bar represents 10 μm in (A–D, F–M) and 25 μm in (O–R). Error bars represent SDs. Student's t-test, ****p < 0.0001. NS, non-significant (p > 0.05)
3.7 Depletion of ClC-c inhibits the maturation of Rab5-labelled early endosomes and Rab7-labelled late endosomesOur results indicated that ClC-c regulates the proliferation of ISCs through the EGFR signalling; however, how it affects this signalling pathway is unclear. Many studies have demonstrated that endocytosis affects the EGFR signalling. Ligand stimulation causes EGFR to internalize and be transported through the endocytic pathway. Therefore, endocytosis not only regulates the rate of EGFR degradation and circulation but also regulates the EGFR-mediated signal transduction.43, 44 By analysing our RNA-seq data, we found that knocking down ClC-c downregulates several endocytosis-related genes, including Rab3, Rabex-5, Rab3-GEF, Rab26 and Rab40 (Figure 7A). These findings strongly suggest that ClC-c might affect the vesicle transport system mediated by RAB family proteins. To test this hypothesis, RAB5-labelled early endosomes and RAB7-labelled late endosomes were visualized in esg+ cells. Under the homeostatic conditions, the RAB5-GFP signal was remarkably lower in ClC-c-depleted group than in the control (Figure 7B–D). There was no difference in the level of the RAB7-GFP signal after knocking down ClC-c, compared with the control level (Figure 7E–G). These results indicate that ClC-c depletion affects the activity of early endosomes instead of late endosomes under homeostatic conditions. BLM treatment for 24 h considerably increased the RAB5-GFP signal compared with the level in the untreated group (Figure 7B,D,H,J). However, knocking down ClC-c prevented this increase (Figure 7I,J). Similar results were observed regarding the RAB7-GFP signal after 24 h of BLM treatment (Figure 7K-M). These results indicate that knocking down ClC-c might inhibit endocytosis.

Knocking down ClC-c inhibits the maturation of Rab5-labelled early endosomes and Rab7-labelled late endosomes. (A) Volcano plots of differentially expressed genes between the ClC-c-depleted Drosophila (esg-Gal4-driven ClC-c RNAi) midguts and control Drosophila (esg-Gal4-driven UAS-lacZ) midguts. (B and C) Immunofluorescence images from the R5 region of the midgut of the control (B, esg-Gal4-driven UAS-RAB5-GFP) or ClC-c-depleted flies (C, esg-Gal4-driven UAS-RAB5-GFP; ClC-c RNAi). (D) Quantification of the RAB5-GFP intensities in (B) and (C). Each dot corresponds to one cell. Gut n ≥ 5; cell n ≥ 50. (E and F) Immunofluorescence images from the R5 region of the midgut of the control (E, esg-Gal4-driven UAS-RAB7-GFP) or ClC-c-depleted (F, esg-Gal4-driven UAS-RAB7-GFP; ClC-c RNAi) flies. (G) Quantification of the RAB7-GFP intensities in (E) and (F). Each dot corresponds to one cell. Gut n ≥ 5; cell n ≥ 50. (H and I) Immunofluorescence images from the R5 region of the midgut of the control (H, esg-Gal4-driven UAS-RAB5-GFP) or ClC-c-depleted (I, esg-Gal4-driven UAS-RAB5-GFP; ClC-c RNAi) flies with BLM treatment. (J) Quantification of the RAB5-GFP intensities in (H) and (I). Each dot corresponds to one cell. Gut n ≥ 10; cell n ≥ 50. (K and L) Immunofluorescence images from the R5 region of the midgut of the control (K, esg-Gal4-driven UAS-RAB7-GFP) or ClC-c-depleted (L, esg-Gal4-driven UAS-RAB7-GFP; ClC-c RNAi
留言 (0)