
記住我

Bacteria play a critical role in influencing the vaginal environment in terms of biochemical and inflammatory properties. Therefore, the characteristics of the VMB may affect conception and the ability to carry a fetus to term. Considering that hormonal changes are associated with significant changes in vaginal microbiome (VMB, see part I), one might expect changes in VMB during pregnancy, reflecting physiologic changes linked to the elevated levels of estrogen and progesterone. In addition, infection has long been recognized as a risk factor for poor reproductive outcomes.1 Dysbiosis, and specifically bacterial vaginosis (BV), is associated with various pregnancy complications,2 including preterm premature rupture of membranes (PPROMs), spontaneous preterm labor, and preterm birth (PTB; see hereinafter), indicating the possible role of VMB in maintenance of normal pregnancy and reproductive outcomes. A better understanding of the dynamics of the VMB during pregnancy, and the possible relations between the VMB and reproduction, may lead to better diagnostic tools and treatments for complications associated with the complex processes of conception, pregnancy, and birth.
Genital Tract Microbiome and InfertilityInfertility, defined as the inability to conceive after 1 year of regular unprotected intercourse, affects 1 in 7 couples.3 Female factors, responsible for 35%–40% of cases, include hormonal changes, tubal occlusion, uterine pathologies, maternal age, and systemic or genetic diseases. However, in approximately 35% of couples, the causes remain unexplained despite a thorough assessment,3 defined as “idiopathic.” Assisted reproductive techniques (ARTs), including in vitro fertilization (IVF), are being offered based on the diagnosis; however, the percentage of implantation and pregnancy rates per embryo transfer remains low. The challenge of improving these outcomes is still an ongoing one.
The female reproductive tract microbiome has been proposed to affect infertility and the success of ART. Different microbial-associated mechanisms were posited as contributing,3 including: pelvic inflammatory disease associated with the presence of specific bacteria in the uterus, tubal occlusion, the correlation between certain bacteria and endometriosis,4 the composition of endometrial microbiome, and the entrance of cervicovaginal bacteria to the upper genital tract with the sperm during fertilization. However, because of the difficulties in obtaining upper reproductive system samples from healthy women, few data are available to date.
Increasing evidence has shown the presence of microorganisms not only in the vagina but also in the uterus, which was previously considered a “sterile niche.”5–7 Lactobacilli are the most represented genus in the endometrium according to some studies6,8,9 but not all10; however, it is estimated that the upper reproductive system hosts 10,000 times fewer bacteria than the vagina.11 The uterine microbiota has been proposed to be able to modulate the endometrial cells' functions as well as the local immune system and to prevent uterine infections.
It has been suggested that infertile women host a different microbiota, both in the lower and/or the upper reproductive tract, compared with fertile women.12–15 Moreno et al.9 compared the endometrial and VMB of fertile, healthy women and found similar microbiome profiles between the 2 analyzed body sites in 80% of the cases. Campisciano et al.14 found significant variations of VMB in idiopathic infertile women compared with 3 groups: healthy, BV-affected, and nonidiopathic infertile women, distinguishing the idiopathic infertile women from the others. Wee et al.13 compared fertile and infertile women and found that Ureaplasma and Gardnerella were more abundant, respectively, in the vagina and the cervix of the latter.
Amato et al.16 analyzed the vaginal and seminal microbiome of 23 couples with idiopathic infertility undergoing intrauterine insemination, aiming to correlate microbial features with pregnancy rates. They found that VMB of idiopathic infertile women differed from controls, as well as different patterns of Lactobacillus species among idiopathic infertile women, with L. crispatus being associated with higher rate of intrauterine insemination success. Therefore, they suggested including VMB evaluation in the work-up of idiopathic infertility, before recommending intrauterine insemination.16
In addition, it has been suggested that the outcomes of IVF procedures may be affected by the resident microbiota. While lactobacilli have been reported to exert beneficial effects, endometrial or vaginal dysbiosis has been related to a worse success rate in terms of lower implantation rates, pregnancy rates, ongoing pregnancies, and live birth rates.9,17
In summary, current data suggest that microbial dysbiosis is associated with infertility and may also play a limited role in IVF outcomes.18 The uterine microbiota, because of its low biomass and the difficult sampling, is challenging to be studied. It also seems that the evaluation of the partner's semen microbiota composition is important as it has implications not only on the composition of the female one but also on the reproductive health of the couple and offspring.3
Vaginal Microbiome and Pregnancy OutcomesAccumulating evidence links microorganisms to the etiology of various maternal-fetal conditions, including PTB, PPROM, fetal growth restriction, late abortions, and stillbirth.1 Preterm birth, defined as birth before 37th gestational weeks, is the leading cause of neonatal morbidity and mortality worldwide, with an estimated 15 million babies throughout the world born preterm, and approximately 1 million of them dying from complications of PTB.1 Several cohort studies have shown that healthy women with normal pregnancy outcomes generally maintain a VMB dominated by Lactobacillus species throughout the entire pregnancy.19
How the VMB may promote healthy pregnancy outcomes is still under investigation.20 However, cultivating bacteria has not revealed the precise etiologies of PTB or other adverse obstetric conditions. As it is estimated that 90% of the microorganisms comprising human microbiomes are uncultivable,21 the development of DNA sequencing technologies may assist in understanding the relationships between the microbiome and pregnancy outcomes.
The Vaginal Microbiome in PregnancyHuman pregnancy is characterized by an increase of lactobacilli abundance in the vaginal milieu, as well as stable vaginal bacterial composition and low bacterial diversity.22
Hormones probably play a crucial factor in determining the VMB composition during pregnancy. Placental estrogen production in pregnancy significantly increases its levels, stimulating maturation of the vaginal epithelium and promoting additional glycogen production and lactobacilli thrive. α-Amylase from the vaginal mucosa processes glycogen to produce maltose, maltotriose, and maltotetraose, which all contribute to lactobacilli growth.23
The shift toward Lactobacillus species dominance occurs early in pregnancy and is most dramatically observed in women of African ancestry. Recently, Serrano et al.24 showed that among women who experienced uncomplicated term birth, the VMB changed during pregnancy, becoming more lactobacilli dominated at the expense of Gardnerella vaginalis and other anaerobes. Stratification of the profiles according to ancestry revealed minimal differences in the microbiome profiles of pregnant and nonpregnant women of European ancestry. However, pregnant women of African and Hispanic ancestry showed a higher prevalence of bacterial communities dominated by Lactobacillus species than their nonpregnant counterparts. The alpha diversities (the diversity within a sample) of the VMB of pregnant women in all groups were lower than those of their nonpregnant counterparts. These findings are in agreement with previous studies.25,26
Together, these observations suggest that the compositions of the VMB of women of African and non-African ancestry respond differently during pregnancy due to complex interactions between human and microbial physiology as well as environmental influences, implying that in the case of a high-diversity VMB, the composition could change during pregnancy, reducing adverse obstetrical risks.
MacIntyre et al.27 reported that a specific lactobacilli community was not required for a term uncomplicated birth. Inline, the predominant Lactobacillus species during pregnancy differs according to various studies26,28–30; in the cohort of MacIntyre et al.,27 the predominant species was L. crispatus, but Asian and White women were also colonized by Lactobacillus jensenii or Lactobacillus gasseri.27 Others have reported that Lactobacillus iners decreased in the second and third trimester compared with the first trimester, whereas L. crispatus increased in the second trimester.28 Nevertheless, in other cohorts, the VMB was mostly dominated by L. iners.26
There is a complex interplay between the VMB, local immune response, and its metabolic milieu.31 Lactic acid increases the release of interleukin (IL)-1β and IL-8 from vaginal epithelial cells. This suggests a synergistic association flanked by inflammatory activation in the host and microbial composition. These activities are most probably dependent on mutually intrinsic (genetic) and extrinsic (environmental) factors. It is known that vaginal epithelial cells only produce the l-lactic acid isomer. Instead, lactobacilli and other lactic acid–producing bacteria produce both the d- and the l-lactic acid isomers. Witkin et al.32 discovered elevated levels of d-lactic acid in vaginas that were colonized almost exclusively by L. crispatus. They suggested that an increase in the proportion of d- to l-lactic acid promoted the expression of metalloproteinase inducer in control of activating the matrix MP-8 and consequently modify the integrity of the uterine cervix responsible for cervical ripening.23,32
Vaginal Microbiome and MiscarriagesMiscarriages, complicating up to 25% of pregnancies, result from various factors, in half of the cases from chromosomal aberrations. However, evidence supports an infectious etiology as well, with reported histological chorioamnionitis in 77% of miscarriage samples compared with 0% of control cases of induced loss for fetal anomaly.33 Furthermore, a relation between BV and increased risk of miscarriage was reported.34
In a study aimed to characterize vaginal bacterial composition in early pregnancy and its relationship with miscarriages, it was found that first trimester miscarriages were associated with Lactobacilli species depletion and a significantly higher proportion of samples dominated by CST IV (see part I).35 Consistent with this, bacterial alpha diversity was significantly higher in miscarriage samples than in matched control samples.
The certain mechanism associating first trimester miscarriages with vaginal dysbiosis is still undetermined. When elucidated, it may represent a modifiable target for miscarriage prevention.35
Vaginal Microbiome and Preterm BirthPreterm birth is considered a multietiological phenomenon, and infection is thought to contribute to at least one third of these cases.19 Preterm birth associated with infection is assumed to be secondary to pathogen ascension from the vagina. This hypothesis is supported by the similarity observed between bacteria found in the placenta and fetal membranes of PTB cases and vaginal microbiota. As a result, studies were conducted aiming to characterize VMB in women with PTB or PPROM. This topic was recently reviewed by Bayar et al.,19 which concluded that the most consistent finding across almost all studies is the benefit of a vaginal microbiota dominated by L. crispatus.
In the United States, women of African ancestry are significantly more likely than women of European ancestry to experience a PTB and PPROM. The difference is presumably attributable to multiple factors, including environmental factors, socioeconomic status, and genetic factors.24 Nevertheless, the concept that ethnicity is a strong determinant of the VMB and its effect on PTB rates has been extensively studied. In a study including mostly White population, Lactobacillus-depleted vaginal CST were correlated with reduced gestational age at delivery, and high abundances of Gardnerella or Ureaplasma were associated with greater risk for PTB.36 A subsequent study showed that although Lactobacillus species depletion and greater abundance of Gardnerella were more common in women of African ancestry, it represented a risk factor for PTB only in White women.37 As part of the integrative Human Microbiome Project, Fettweis et al.38 compared the VMB and cytokine profile between women of African ancestry delivering preterm and at term. Analyses showed that the former had significantly lower levels of L. crispatus, higher levels of BVAB1, Sneathia amnii, TM7-HI, and a group of Prevtolla species. Women who delivered at term were more likely to have L. crispatus and reduced prevalence of Atopobium vaginae and G. vaginalis. Preterm birth was also associated with a vaginal cytokine profile richer in proinflammatory cytokines, including eotaxin, IL-1β, IL-6, and MIP-1β. In European studies comprising mostly White women,19 an increased risk of PTB was associated with L. iners, whereas a protective effect for L. crispatus dominance was found.39,40 Other studies showed that vaginal dysbiosis is associated with PPROM and constitutes a risk factor for chorioamnionitis as well as for neonatal sepsis.19
SummarySeveral of the negative pregnancy outcomes have been associated with an altered vaginal milieu. Dysbiosis is remarkably linked with poor pregnancy outcomes from preconception to delivery. More research is essential to explore the mechanisms causing adverse events and the relationship between the VMB and the immune system. The most consistent finding across almost all studies is the benefit of a vaginal microbiota dominated by L. crispatus. Many studies have examined the efficacy of antibiotics for the treatment and prevention of PTB, and almost entirely targeted BV. The failure of antibiotic therapy to improve PTB in the context of abnormal VMB, has led to an interest in the modulation of the VMB using live biotherapeutic products or probiotics (see part V). To date, none of the treatment modalities that were studied showed beneficial effects upon the PTB rate.19
THE VAGINAL MICROBIOME AND GYNECOLOGIC CANCERS IntroductionMicroorganisms, including viruses and bacteria, are estimated to have a role in 15% of malignant neoplasms.41 Oncogenic bacteria and viruses directly modulate carcinogenesis through specific toxins that can damage host DNA or by integration of oncogenes into host genomes, respectively. Nevertheless, clinical and animal studies suggest that carcinogenesis may be driven by global changes in the microbiome, rather than by single pathogens.42 In this case, the pathophysiology is believed to result from altered host defense responses to dysbiotic microbiota.43 Dysbiosis may promote epithelial barrier dysfunction, immune dysregulation, genotoxicity, and/or inflammation, collectively creating a tumor permissive microenvironment.42–44 Interestingly, some bacteria, such as Chlamydia trachomatis, induce epithelial-to-mesenchymal transition of infected cells, which might promote loss of epithelial cell adhesion and downregulation of DNA damage responses, inducing carcinogenesis.45 In addition, bacterial communities may contribute to the etiology, disease severity, and/or response to treatment of gynecological malignancies.46
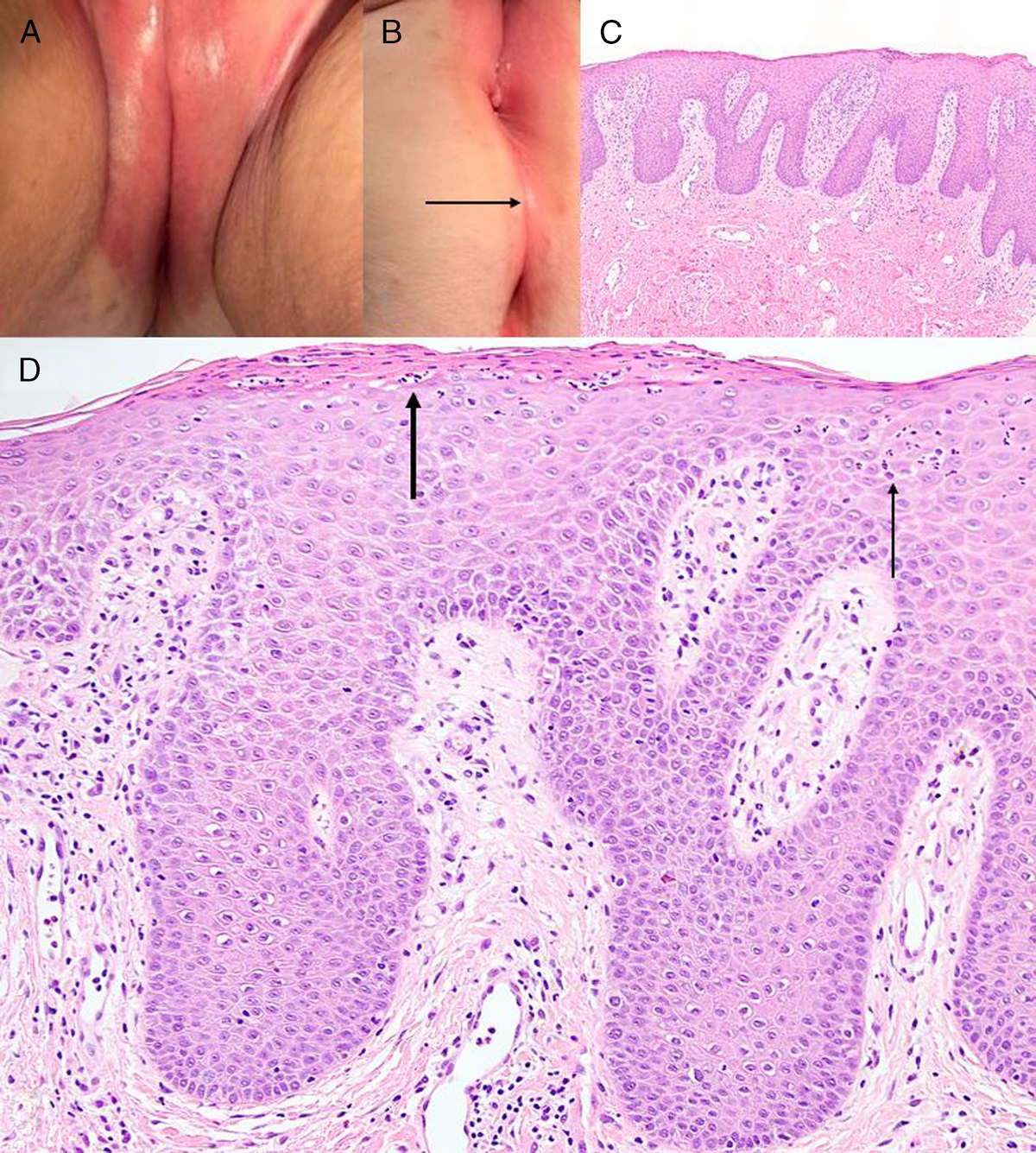
Cervical Cancer, the Vaginal Microbiome, and Human PapillomavirusCervical carcinoma (CC) is the most common human papillomavirus (HPV)–related malignancy, with high-risk HPV genotypes, especially HPV-16 and HPV-18, being well-recognized oncogenic factors, detected in 99.7% of the cases.47 Of high-risk HPV infections, 85%–90% are spontaneously cleared and only 10%–15% persist, consequently causing precancerous cervical intraepithelial neoplasia. These lesions may subsequently progress to invasive CC.48
Various factors, including age of sexual debut, parity, usage of contraceptives, smoking, and other STIs, have been shown to increase the risk of progression to cervical neoplasia among HPV-infected women.49–52 Emerging evidence suggests that the VMB also participates in cervical carcinogenesis.52–55 Studies reported associations between non-Lactobacillus–dominant VMB, HPV infection, and its persistence.52–55 In addition, BV has been associated with an increased risk of HPV acquisition and decreased clearance.56
Three meta-analyses supported a link between a non-Lactobacillus–dominant VMB and cervical disease via the effect of the microbiome on HPV acquisition, persistence, and the development of cervical intraepithelial neoplasia (CIN).57–59 Vaginal microbiome dominated by L. iners was also associated with higher odds of HPV and progression to cervical neoplasia compared with L. crispatus dominance.57
Three microbiome studies have included women with cervical abnormalities and cancer.53–55 They consistently reported depletion of Lactobacilli species and a substantial increase in VMB diversity in women with cervical intraepithelial neoplasia and invasive carcinoma compared with healthy women. Aiming at identifying particular bacteria, which are associated with cervical disease, they pointed out that 3 BV-associated microorganisms: Sneathia sanguinegens, Anaerococcus tetradius, and Peptostreptococcus anaerobius were considerably more abundant among patients with high-grade CIN than in those with low-grade abnormalities.53 The presence of Sneathia in the vagina was suggested as a metagenomic marker for HPV persistence and progression of CIN.52 Not unexpectedly, a significant increase in vaginal pH, correlated with the depletion of lactobacilli, was related to the severity of cervical disease progression.55
The increasing body of evidence implies a complex relationship between the host, local microbiome, and HPV during cervical carcinogenesis. A cross-sectional study evaluated multiple immune mediators in the cervicovaginal microenvironment in women with CC/CIN and those without neoplasia (HPV infected or not). Nonlactobacilli-dominated VMB was associated with increased proinflammatory and chemotactic mediators,55 as well as with altered local metabolic profiles,60 which might directly or indirectly contribute to cervical carcinogenesis.61
Endometrial Cancer and Vaginal MicrobiomeEnvironmental factors, including obesity, unopposed estrogen exposure, and inflammation, are considered major risk factors for endometrial cancer (EC) development.62,63
Endometrial colonization with bacteria was hypothesized to promote carcinogenesis via microbiota-mediated stimulation of cytokines secretion from the host cells or by mediating dysbiosis-related growth factors.42,64
Walther-António et al.65 studied the microbiome in patients with endometrial hyperplasia, EC, and benign uterine conditions, using samples that were taken from various sites along the reproductive tract. The microbiome sequencing revealed that the microbiomes of the vagina, cervix, fallopian tubes, and ovaries are significantly correlated. There was a structural microbiome shift in the EC and hyperplasia cases, distinguishable from benign cases. In samples belonging to the EC cohort, several taxa were found to be significantly increased, including A. vaginae and Porphyromonas species, which were found to be associated with disease status, especially if combined with a vaginal pH >4.5. The authors suggested that a possible subclinical vaginal infection might cause chronic upper genital tract infection and inflammation that may trigger carcinogenesis.66
Another hypothesis correlates the associations between intestinal and VMBs as related with EC.46 Gut microbiome composition can affect the risk of EC directly by increasing adiposity, with a resultant increase in circulating estrogens, or by indirectly governing the metabolism of estrogens.61,66,67
The gut microbiota can indirectly influence the genital microbiota composition through estrogen-mediated mechanisms.52,67,68 Estrogen facilitates the growth of lactobacilli through induction of glycogen production in epithelial cells; however, it was shown that circulating estrogen levels are influenced by gut microbiota,69 leading to a concept of an estrogen-mediated gut-vagina axis.67 The relationship between these 2 mucosal sites involves enteric bacteria that metabolize estrogens; the collection of these microorganisms and their genes was termed the “estrobolome.”68 These microorganisms secrete enzymes (β-glucuronidase and β-glucosidase), which deconjugate hepatically conjugated estrogens, resulting in free estrogen, which is absorbed to the circulation,69 subsequently transported to distal sites, where it binds to its receptors and produces its physiological functions. Thus, an alteration of the gut microbiota diversity, which may result in a lack of estrogen-metabolizing bacteria could influence the VMB composition via the estrobolome. Similarly, a dysbiotic gut microbiome that increases estrogens deconjugation will be followed by increased estrogen levels and hyperestrogenic environment, which in turn increase the risk of estrogen-dependent tumors, including EC.70
Microbiota in Tubal and Ovarian CancerGenital dysbiosis has been associated with the development of ovarian carcinoma (OC).71,72 Two cross-sectional studies have compared ovarian tissues in women with OC and healthy women and found a distinct ovarian microbiome in women with OC.71,72 Specifically, the presence of potentially pathogenic intracellular microorganisms, such as Brucella, Mycoplasma, and Chlamydia species, was found in 60%–76% of ovarian tumors.72–74
Other pathogenic microorganisms that were identified in OC tissues included HPV, cytomegalovirus, and C. trachomatis.75 Moreover, the microbiome of the malignant ovarian tissue had distinct microbial signatures compared with the healthy surrounding ovarian tissues within the same individuals.72 The authors concluded that OC tissue has a significantly different microbiome composition compared with that of surrounding tissue and controls.72
Although these studies suggested a link with inflammation, the link between microbiota and ovarian cancer remains unclear. These microorganisms might induce carcinogenesis through direct or indirect mechanisms; however, it is also possible that the tumor microenvironment might also favor the recruitment and growth of anaerobic microorganisms.
The fallopian tubes and ovaries are low-abundancy sites, which makes sample collection without cross-contamination and interpretation of the results challenging.52 Infertility is considered an independent risk factor for OC. The causes of female infertility are variable, and 14% of the overall causes of infertility are linked to tubal factors. These are usually resulting from salpingitis caused by a previous infection, such as pelvic inflammatory disease.66 Infections, leading to infertility caused by “tubal factors,” may also cause subclinical and possibly persistent microbial infection. With recent advances in the understanding of OC, it is now thought that the fimbriae portion of the fallopian tube is the origin of OC. Subclinical microbial infections within the fallopian tube may cause both infertility and OC.66
SummaryThe associations between the microbiome and gynecologic cancers are largely unknown. Proving causation in cancer is difficult because of the complex interactive nature of potential causative factors. These difficulties can be overcome by conducting large, international, longitudinal cohort studies.76 The female genital tract may be more susceptible to microbial insults because it is exposed to the external environment and is populated with numerous bacterial species, like the gut. Certain elements of the microbiota have been shown to provoke inflammatory reactions, whereas others produce anti-inflammatory reactions; this balance might be impaired with a change in microbial variety.
REFERENCES 1. Solt I. The human microbiome and the great obstetrical syndromes: a new frontier in maternal-fetal medicine. Best Pract Res Clin Obstet Gynaecol 2015;29:165–75. 2. Lamont RF, Sobel JD, Akins RA, et al. The vaginal microbiome: new information about genital tract flora using molecular based techniques. BJOG 2011;118:533–49. 3. Tomaiuolo R, Veneruso I, Cariati F, et al. Microbiota and human reproduction: the case of male infertility. High Throughput 2020;9:10. 4. Leonardi M, Hicks C, El-Assaad F, et al. Endometriosis and the microbiome: a systematic review. BJOG 2020;127:239–49. 5. Franasiak JM, Scott RT. Reproductive tract microbiome in assisted reproductive technologies. Fertil Steril 2015;104:1364–71. 6. Mitchell CM, Haick A, Nkwopara E, et al. Colonization of the upper genital tract by vaginal bacterial species in nonpregnant women. Am J Obstet Gynecol 2015;212:611.e1–9. 7. Moreno I, Franasiak JM. Endometrial microbiota—new player in town. Fertil Steril 2017;108:32–9. 8. Franasiak JM, Werner MD, Juneau CR, et al. Endometrial microbiome at the time of embryo transfer: next-generation sequencing of the 16S ribosomal subunit. J Assist Reprod Genet 2016;33:129–36. 9. Moreno I, Codoñer FM, Vilella F, et al. Evidence that the endometrial microbiota has an effect on implantation success or failure. Am J Obstet Gynecol 2016;215:684–703. 10. Verstraelen H, Vilchez-Vargas R, Desimpel F, et al. Characterisation of the human uterine microbiome in non-pregnant women through deep sequencing of the V1-2 region of the 16S rRNA gene. PeerJ 2016;2016:e1602. 11. Chen C, Song X, Wei W, et al. The microbiota continuum along the female reproductive tract and its relation to uterine-related diseases. Nat Commun 2017;8:875. 12. Moreno I, Simon C. Deciphering the effect of reproductive tract microbiota on human reproduction. Reprod Med Biol 2019;18:40–50. 13. Wee BA, Thomas M, Sweeney EL, et al. A retrospective pilot study to determine whether the reproductive tract microbiota differs between women with a history of infertility and fertile women. Aust N Z J Obstet Gynaecol 2018;58:341–8. 14. Campisciano G, Florian F, D'Eustacchio A, et al. Subclinical alteration of the cervical-vaginal microbiome in women with idiopathic infertility. J Cell Physiol 2017;232:1681–8. 15. Moreno I, Simon C. Relevance of assessing the uterine microbiota in infertility. Fertil Steril 2018;110:337–43. 16. Amato V, Papaleo E, Pasciuta R, et al. Differential composition of vaginal microbiome, but not of seminal microbiome, is associated with successful intrauterine insemination in couples with idiopathic infertility: a prospective observational study. Open Forum Infect Dis 2020;7:ofz525. 17. Haahr T, Jensen JS, Thomsen L, et al. Abnormal vaginal microbiota may be associated with poor reproductive outcomes: a prospective study in IVF patients. Hum Reprod 2016;31:795–803. 18. Karaer A, Doğan B, Günal S, et al. The vaginal microbiota composition of women undergoing assisted reproduction: a prospective cohort study. BJOG 2021. doi:10.1111/1471-0528.16782. 19. Bayar E, Bennett PR, Chan D, et al. The pregnancy microbiome and preterm birth. Semin Immunopathol 2020;42:487–99. 20. Gandhi K, Gutierrez P, Garza J, et al. Vaginal Lactobacillus species and inflammatory biomarkers in pregnancy. Minerva ginecol 2020;72:299–309. 21. Dethlefsen L, McFall-Ngai M, Relman DA. An ecological and evolutionary perspective on human-microbe mutualism and disease. Nature 2007;449:811–8. 22. Gupta S, Kakkar V, Bhushan I. Crosstalk between vaginal microbiome and female health: a review. Microb Pathog 2019;136:103696. 23. Amabebe E, Anumba DOC. The vaginal microenvironment: the physiologic role of Lactobacilli. Front Med (Lausanne) 2018;5:181. 24. Serrano MG, Parikh HI, Brooks JP, et al. Racioethnic diversity in the dynamics of the vaginal microbiome during pregnancy. Nat Med 2019;25:1001–11. 25. Aagaard K, Riehle K, Ma J, et al. A metagenomic approach to characterization of the vaginal microbiome signature in pregnancy. PLoS One 2012;7:e36466. 26. Romero R, Hassan SS, Gajer P, et al. The composition and stability of the vaginal microbiota of normal pregnant women is different from that of non-pregnant women. Microbiome 2014;2:4. 27. MacIntyre DA, Chandiramani M, Lee YS, et al. The vaginal microbiome during pregnancy and the postpartum period in a European population. Sci Rep 2015;5:8988. 28. Zheng N, Guo R, Yao Y, et al. Lactobacillus iners is associated with vaginal dysbiosis in healthy pregnant women: a preliminary study. Biomed Res Int 2019;2019:6079734. 29. Hyman RW, Fukushima M, Jiang H, et al. Diversity of the vaginal microbiome correlates with preterm birth. Reprod Sci 2014;21:32–40. 30. Human Microbiome Project Consortium. Structure, function and diversity of the healthy human microbiome. Nature 2012;486:207–14. 31. Linhares IM, Sisti G, Minis E, et al. Contribution of epithelial cells to defense mechanisms in the human vagina. Current Infectious Disease Reports 2019;21. 32. Witkin SS, Moron AF, Ridenhour BJ, et al. Vaginal biomarkers that predict cervical length and dominant bacteria in the vaginal microbiomes of pregnant women. mBio 2019;10:e02242. 33. Allanson B, Jennings B, Jacques A, et al. Infection and fetal loss in the mid-second trimester of pregnancy. Aust N Z J Obstet Gynaecol 2010;50:221–5. 34. Hay PE, Lamont RF, Taylor-Robinson D, et al. Abnormal bacterial colonisation of the genital tract and subsequent preterm delivery and late miscarriage. BMJ 1994;308:295–8. 35. Al-Memar M, Bobdiwala S, Fourie H, et al. The association between vaginal bacterial composition and miscarriage: a nested case-control study. BJOG 2020;127:264–74. 36. DiGiulio DB, Callahan BJ, McMurdie PJ, et al. Temporal and spatial variation of the human microbiota during pregnancy. Proc Natl Acad Sci U S A 2015;112:11060–5. 37. Callahan BJ, DiGiulio DB, Aliaga Goltsman DS, et al. Replication and refinement of a vaginal microbial signature of preterm birth in two racially distinct cohorts of US women. Proc Natl Acad Sci U S A 2017;114:9966–71. 38. Fettweis JM, Serrano MG, Brooks JP, et al. The vaginal microbiome and preterm birth. Nat Med 2019;25:1012–21. 39. Petricevic L, Domig KJ, Nierscher FJ, et al. Characterisation of the vaginal Lactobacillus microbiota associated with preterm delivery. Sci Rep 2014;4:5136. 40. Kindinger LM, Bennett PR, Lee YS, et al. The interaction between vaginal microbiota, cervical length, and vaginal progesterone treatment for preterm birth risk. Microbiome 2017;5:6. 41. Shahanavaj K, Gil-Bazo I, Castiglia M, et al. Cancer and the microbiome: potential applications as new tumor biomarker. Expert Rev Anticancer Ther 2015;15:317–30. 42. Schwabe RF, Jobin C. The microbiome and cancer. Nat Rev Cancer 2013;13:800–12. 43. Garrett WS. Cancer and the microbiota. Science 2015;348:80–6. 44. Rajagopala SV, Vashee S, Oldfield LM, et al. The human microbiome and cancer. Cancer Prev Res 2017;10:226–34. 45. Zadora PK, Chumduri C, Imami K, et al. Integrated phosphoproteome and transcriptome analysis reveals chlamydia-induced epithelial-to-mesenchymal transition in host cells. Cell Reports 2019;26:1286–1302.e8. 46. Chase D, Goulder A, Zenhausern F, et al. The vaginal and gastrointestinal microbiomes in gynecologic cancers: a review of applications in etiology, symptoms and treatment. Gynecol Oncol 2015;138:190–200. 47. Walboomers JM, Jacobs MV, Manos MM, et al. Human papillomavirus is a necessary cause of invasive cervical cancer worldwide. J Pathol 1999;189:12–9. 48. Shulzhenko N, Lyng H, Sanson GF, et al. Ménage à trois: an evolutionary interplay between human papillomavirus, a tumor, and a woman. Trends Microbiol 2014;22:345–53. 49. Gravitt PE, Winer RL. Natural history of HPV infection across the lifespan: role of viral latency. Viruses 2017;9:267. 50. Ryser MD, Rositch A, Gravitt PE. Modeling of US human papillomavirus (HPV) seroprevalence by age and sexual behavior indicates an increasing trend of HPV infection following the sexual revolution. J Infect Dis 2017;216:604–11. 51. Mhatre M, McAndrew T, Carpenter C, et al. Cervical intraepithelial neoplasia is associated with genital tract mucosal inflammation. Sex Transm Dis 2012;39:591–7. 52. Łaniewski P, Ilhan ZE, Herbst-Kralovetz MM. The microbiome and gynaecological cancer development, prevention and therapy. Nat Rev Urol 2020;17:19–22. 53. Mitra A, MacIntyre DA, Lee YS, et al. Cervical intraepithelial neoplasia disease progression is associated with increased vaginal microbiome diversity. Sci Rep 2015;5:1–11. 54. Audirac-Chalifour A, Torres-Poveda K, Bahena-Román M, et al. Cervical microbiome and cytokine profile at various stages of cervical cancer: A pilot study. PLoS One 2016;11:e0153274. 55. Łaniewski P, Barnes D, Goulder A, et al. Linking cervicovaginal immune signatures, HPV and microbiota composition in cervical carcinogenesis in non-Hispanic and Hispanic women. Scientific Reports 2018;8:7593. 56. Watts DH, Fazarri M, Minkoff H, et al. Effects of bacterial vaginosis and other genital infections on the natural history of human papillomavirus infection in HIV-1-infected and high-risk HIV-1-uninfected women. J Infect Dis 2005;191:1129–39. 57. Norenhag J, Du J, Olovsson M, et al. The vaginal microbiota, human papillomavirus and cervical dysplasia: a systematic review and network meta-analysis. BJOG 2020;127:171–80. 58. Brusselaers N, Shrestha S, van de Wijgert J, et al. Vaginal dysbiosis and the risk of human papillomavirus and cervical cancer: systematic review and meta-analysis. Am J Obstet Gynecol 2019;221:9–18.e8. 59. Wang H, Ma Y, Li R, et al. Associations of cervicovaginal lactobacilli with high-risk human papillomavirus infection, cervical intraepithelial neoplasia, and cancer: a systematic review and meta-analysis. J Infect Dis 2019;220:1243–54. 60. Srinivasan S, Morgan MT, Fiedler TL, et al. Metabolic signatures of bacterial vaginosis. mBio 2015;6:1–16. 61. Łaniewski P, Cui H, Roe DJ, et al. Features of the cervicovaginal microenvironment drive cancer biomarker signatures in patients across cervical carcinogenesis. Sci Rep 2019;9:7333. 62. Allen NE, Key TJ, Dossus L, et al. Endogenous sex hormones and endometrial cancer risk in women in the European Prospective Investigation into Cancer and Nutrition (EPIC). Endocr Relat Cancer 2008;15:485–97. 63. Dossus L, Rinaldi S, Becker S, et al. Obesity, inflammatory markers, and endometrial cancer risk: a prospective case-control study. Endocr Relat Cancer 2010;17:1007–19. 64. Baker JM, Chase DM, Herbst-Kralovetz MM. Uterine microbiota: residents, tourists, or invaders? Front Immunol 2018;9:208. 65. Walther-António MRS, Chen J, Multinu F, et al. Potential contribution of the uterine microbiome in the development of endometrial cancer. Genome Med 2016;8:1–15. 66. Mert I, Walther-Antonio M, Mariani A. Case for a role of the microbiome in gynecologic cancers: clinician's perspective. J Obstet Gynaecol Res 2018;44:1693–704. 67. Baker JM, Al-Nakkash L, Herbst-Kralovetz MM. Estrogen-gut microbiome axis: physiological and clinical implications. Maturitas 2017;103:45–53. 68. Plottel CS, Blaser MJ. Microbiome and malignancy. Cell Host Microbe 2011;10:324–35. 69. Flores R, Shi J, Fuhrman B, et al. Fecal microbial determinants of fecal and systemic estrogens and estrogen metabolites: a cross-sectional study. J Transl Med 2012;10:253. 70. Chen KL, Madak-Erdogan Z. Estrogen and microbiota crosstalk: should we pay attention? Trends Endocrinol Metab 2016;27:752–5. 71. Zhou B, Sun C, Huang J, et al. The biodiversity composition of microbiome in ovarian carcinoma patients. Sci Rep 2019;9:1691. 72. Banerjee S, Tian T, Wei Z, et al. The ovarian cancer oncobiome. Oncotarget 2017;8:36225–45. 73. Chan PJ, Seraj IM, Kalugdan TH, et al. Prevalence of mycoplasma conserved DNA in malignant ovarian cancer detected using sensitive PCR-ELISA. Gynecol Oncol 1996;63:258–60. 74. Emara M, Vyas V, al Awadi S, et al. Synchronous occurrence of brucellosis and ovarian cancer—a case report. Austral-Asian J Cancer 2007;6:257–9. 75. Shanmughapriya S, Vasanthi N, SenthilKumar G, et al. Viral and bacterial aetiologies of epith
留言 (0)