Stroke can occur because of blockage (ischaemic stroke) or rupture (haemorrhagic stroke) of cerebral blood vessels and is a serious neurological disorder that results in long-term sequelae if left untreated. Restoring cerebral flow and minimizing neuronal loss are two major aims of acute stroke therapy. Over the last two decades, therapeutic strategies have included intravenous thrombolysis and mechanical thrombectomy, triggering a revolution in reperfusion therapy for acute stroke. Recent advances in neuroscience and molecular biology have provided novel insights into how neuronal energy metabolism is affected post-stroke. Physiological energy homeostasis drives neural network dynamics which maintain normal brain function. In clinical practice, modern neuroimaging techniques have unlocked new avenues for the selection of patients that can receive more targeted therapies, which may help to restore the metabolism homeostasis in limited time and harsh microenvironments for decreasing neuronal deterioration following a stroke. This remarkable progress has improved our mechanistic understanding of stroke and created new opportunities for developing innovative therapeutic strategies targeting the energy metabolism of neurons.
In the current special issue of the Journal of Neurochemistry, entitled “Stroke and Energy Metabolism” (https://onlinelibrary.wiley.com/doi/toc/10.1111/(ISSN)1471-4159.stroke-and-energy), we present three original research articles and six review articles, which cover key aspects of energy metabolism changes following stroke. These articles focus on disturbed cerebral blood flow circulation, mitochondrial dysfunction, programmed neuronal cell death and cell–cell communication relevant to brain energy metabolism. This series of articles provides novel mechanistic insights and lays the foundation for further research as well as effective clinical treatment of stroke patients.
Stroke can be classified into different types depending on the causes and clinical symptoms. For instance subarachnoid haemorrhage mainly results from a ruptured intracranial aneurysm, but most patients with subarachnoid haemorrhage do not exhibit obvious ischaemic lesions or formation of haematoma in the brain. Accordingly, subarachnoid haemorrhage is an ideal model to investigate the association between disturbed intracerebral circulation and brain dysfunction after an acute central nervous system injury. In their article, Zhou and colleagues summarize the three steps of microcirculation disturbance after subarachnoid haemorrhage (Zhou et al., 2021), as well as pivotal pathophysiological processes of intracerebral circulation, including, but not limited to, decreased cerebral blood perfusion, abnormal vascular reactivity and microthrombosis.
Based on the concept of a ‘vascular neural network’ underlying both ischaemic and haemorrhagic stroke, hypoxia because of disturbed intracranial circulation is considered to be the most common and key mechanism for secondary brain injury following stroke (Zhang et al., 2012). Despite the controversy regarding the best course of action and potential hypertransfusion risk in ischaemic-reperfusion injury, successful recovery of intracranial circulation homeostasis is widely considered beneficial for stroke patients. Central sympathetic nerve activation and its modulatory factors are likely to be involved in post-stroke secondary brain injury following subarachnoid haemorrhage and share pathogenic mechanisms with other types of stroke (Hasegawa et al., 2021). Intricated forms of sympathetic modulation may contribute to central and peripheral injury in this condition.
The disturbed intracranial circulation and impaired blood flow to brain tissue following stroke lead to insufficient energy supply to neurons and other intracranial cells. Since mitochondria are the main powerhouse of cells, impaired mitochondrial dysfunction after stroke is of great interest (Chen et al., 2020; Chen et al., 2020). In their article, Yang and colleagues investigate the disruption of mitochondrial transport because of stroke-induced disassembly of microtubules, as well as mitochondrial dysfunction induced by axonal injury after intracerebral haemorrhage. The authors suggest that MEC17-induced α-tubulin acetylation may be a promising strategy to alleviate mitochondrial damage at early stages in order to protect axonal integrity and improve motor control recovery (Yang et al., 2021).
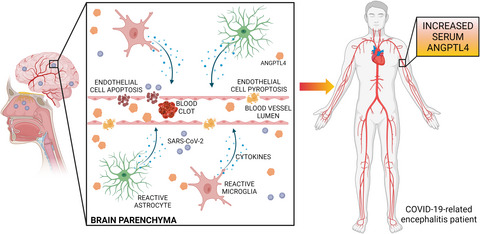
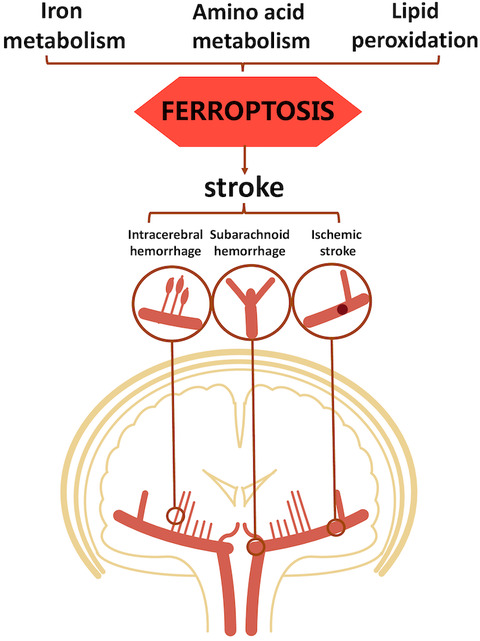
Under prolonged hypoxic stress, mitochondrial dysfunction promotes further neuronal damage and induces various types of programmed neuronal cell death, such as mitophagy (Pickles et al., 2018), ferroptosis (Liu, Fang, et al., 2021) and pyroptosis (He et al., 2021). In their article, Liu and colleagues demonstrate that poly(ADP-ribose) polymerase 1 (PARP-1)-dependent cell death, also referred to as PARthanatos, plays a pivotal role in ischaemic neuronal cell death after stroke (Liu et al., 2021).
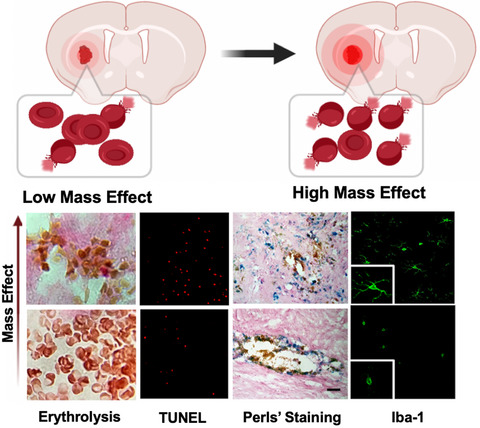
Stroke results in various metabolic issues in injured brain tissue. Multimodal monitoring of a patient's clinical history, including functional neuroimaging and assessment of various metabolic biomarkers may provide critical insight into underlying pathophysiology, with the aim of improved diagnosis, monitoring and treatment of stroke (O'Connell et al., 2020). A practical issue is that the degradation products of erythrolysis (Gong et al., 2021) may mask information derived from markers of brain dysfunction that can be measured in the cerebrospinal fluid, particularly if blood has diffused into the cerebrospinal fluid (Illes, 2017). In this regard, the intracranial glymphatic system and cerebrospinal fluid circulation have been reported to participate in the pathophysiological processes after stroke (Zhou et al., 2021). This special issue includes work by Sands and colleagues presenting evidence for significantly higher levels of systemic Regenerating Family Member 3 Alpha (REG3A) in stroke patients with moderate to severe neuro-functional deficits (Sands et al., 2021). Xu and colleagues summarize the various neurosteroids derived by neuronal or glial mitochondria from cholesterol and other steroidal precursors (Xu et al., 2021). The authors suggest that oestrogen, progesterone, allopregnanolone, dehydroepiandrosterone and vitamin D exert positive therapeutic effects in stroke and its subsequent complications (Xu et al., 2021). In the final article in this special issue, Liddle and colleagues discuss the approach of local hypothermia therapy which, via cooling down of the brain, reduces the metabolic rate of injured brain tissue after stroke, limiting cerebral oxygen and glucose consumption, thus protecting neurons from damage in the acute phase after stroke (Liddle et al., 2021).
Further investigation on mechanisms involved in glia regulation of neuronal death, neuroinflammation and distinct forms of cell–cell communication is warranted to understand new ways to improve brain function (Bonvento & Bolanos, 2021; Kugler et al., 2021). Collectively, this volume discusses the pivotal effects of disturbed intracerebral circulation and energy metabolism after stroke. The progress in proteomics and metabolomics in recent years has enabled comprehensive investigations of the metabolic profiles of various body fluids such as blood and cerebrospinal fluid after stroke, which may unveil pathophysiological mechanisms and therapeutic strategies for stroke. We hope that the current special issue will help to draw attention to the ongoing need for improvements in current stroke treatment strategies, with the aim of minimizing adverse consequences for affected patients.
The author declares no conflict of interest beyond having edited the current special issue “Stroke and Energy Metabolism”.







留言 (0)