
記住我



Fruit morphology and seed (embryonic plant) dispersal mechanisms have been studied for many years (Hughes et al., 1994; Janson, 1983; Pijl, 1969; Ridley, 1930; Tamme et al., 2014; Thomson et al., 2010, among many others). Primary dispersal by frugivorous animals maximizes seed dispersion and distance from the parental tree. Accordingly, the Janzen–Connell hypothesis suggests that increasing space between conspecific seeds should increase their survival by decreasing seed predation and pathogen transmission (Connell, 1970; Janzen, 1970). The Janzen–Connell hypothesis is often used to explain seed dispersal, and the maintenance of diversity in tropical systems where up to 100% of trees rely on animal dispersers (Muller-Landau & Hardesty, 2005; Seidler & Plotkin, 2006). Most studies of seed dispersal and relevant seed traits have focused on primary dispersal by mammals and birds (Bodmer, 1991; Chapman, 1989; Estrada et al., 1993; Galetti et al., 2001; Matías et al., 2010; Milton, 1981). However, primary dispersal alone may have many downsides because seeds are often deposited in an aggregated fashion on the ground and only rarely in an appropriate micro-habitat. This may lead to a decrease in seedling establishment (Vander Wall & Longland, 2004). There are two types of animal seed dispersal, (a) endozoochorous, passing through the digestive system of an animal, and (b) epizoochorous, adhering to an animal usually in their fur. Secondary dispersal of endozoochorous seeds by dung beetles has also been studied but to a lesser degree. There is evidence that seedling survival is higher when both mammals and dung beetles act together as primary and secondary dispersers, respectively (Culot et al., 2018). Understanding secondary dispersal by dung beetles could be critical to our understanding of forest dynamics and seedling establishment because dung beetles reduce seed clumping (Lawson et al., 2012; Urrea-Galeano et al., 2019), which should decrease density-dependent seed mortality (e.g., pathogen and seed predation) (Andresen & Levey, 2004; Beaune et al., 2012; Bell et al., 2006; Vulinec, 2002). When dung beetles bury seeds, they drastically reduce seed discovery by seed predators. In at least one case, burial by dung beetles decreased seed predation rates by as much as 95%–98% (Vulinec, 2002). Primarily dispersed seeds suffer from seed predation accelerated by the fact that dung odors are attractive to seed predators such as rodents (Andresen & Levey, 2004; Beaune et al., 2012). For this reason, seed traits that promote acceptance by dung beetles might be advantageous for seeds.
Studies of seed traits related to the acceptability and burial of seeds by dung beetles have primarily focused only on seed size (Andresen, 2002; Andresen & Levey, 2004; Braga et al., 2017; Feer, 1999). One example of a trait other than size influencing seed acceptability to dung beetles is the odor trait used by Ceratocaryum argenteum, seeds mimic the odor of herbivore feces and are buried by dung beetles. These seeds are protected from seed predation by rodents because dung beetles bury them (Midgley et al., 2015). However, seeds do not provide either the adult or larval dung beetles with sustenance, and as a result, dung beetles have been observed cleaning and removing large seeds from their dung balls. For this reason, it is essential to understand what traits make seeds acceptable to dung beetles.
Small seeds are more likely to be included in dung balls and dispersed by dung beetles. Large beetles accept larger seeds in their dung balls than do smaller beetles (Andresen, 2001). Large seeds are also buried less deeply by dung beetles than small seeds (Shepherd & Chapman, 1998), which is important because seeds that are buried too deeply do not germinate (Andresen & Levey, 2004; Shepherd & Chapman, 1998). Methods for studying seed dispersal by dung beetles focus on size, often excluding other seed traits (e.g., surface or shape). Studies using beads as a proxy for seeds overlook potentially important seed traits, for example, surface or shape (Andresen, 2002; Andresen & Feer, 2005; Braga et al., 2017; Feer, 1999; Lawson et al., 2012).
Dung beetles disperse more small seeds than large seeds, resulting from a combination of rejecting large seeds and a higher abundance of small seeds (Andresen, 2002; Andresen & Feer, 2005; Andresen & Levey, 2004). Large dung beetles move a greater quantity of dung and are therefore expected to disperse more seeds and accept larger seeds than smaller dung beetles (Andresen, 2002; Nichols et al., 2008; Shepherd & Chapman, 1998).
1.1 Hypothesis and predictionsOur field observations of large beetles suggest that they accept larger seeds than smaller beetles. We also made a novel observation that made us consider traits apart from size that might be important in determining a seed's acceptability to a dung beetle. Oxysternon conspicillatum, Canthon angustatus, and Canthidium sp. treated a very large seed Spondias mombin as though it was dung. S. mombin is a large seed covered in hairs (pubescent), and dung sticks to the hairs, possibly misleading the beetles as to its makeup (Figure 1). We formulated a general hypothesis from our field observations: secondary seed dispersal by dung beetles, where advantageous, might be facilitated by some aspects of seed morphology. We focus our analysis on three seed traits: (a) round shape, (b) pubescent surface, and (c) seed size. We tested this hypothesis with the following predictions. (1) Larger dung beetles accept larger seeds in their dung balls, (2) large endozoochorous seeds are rounder, (3) large endozoochorous seed species are more likely to be pubescent than smaller ones, and (4) large pubescent seeds are over-represented in dung balls.

(a) Illustration of Spondias's movement from the spider monkey to the dung beetles Oxysternon conspicillatum, Canthon angustatus, and Canthidium sp. (b) Photograph of an (O. conspicillatum) beetle with the Spondias mombin seed (Ateles fusciceps fusciceps) feces that is pushed into a crevasse, from a brown-headed spider monkey's defecate. (c) Screen grab from a video of C. angustatus and Canthidium sp. trying to use a second S. mombin seed as they normally would use dung
2 METHODS 2.1 Seed traits from in situ collections of dung balls and fecal samplesOur study site is in the Ecuadorian Chocó, a tropical lowland forest on the western side of the Andes within the Reserva Canadé and Tesoro Escondido (00°31′2″N, 79°12′13″W). During our observations of dung beetles in the field from January 2019 to June 2019, we opportunistically collected 82 dung balls from dung beetles = 2, Scybalocanthon trimaculatus N = 1) from under troops of brown-headed spider monkeys (Ateles fusciceps fusciceps) and mantled howler monkeys (Alouatta palliata). We calculated the range of seed sizes for the two common beetle species. Tunneling species O. conspicillatum and S. noctis had more individual balls (N = 33), but they only came from 11 individual beetles. We also collected 29 fecal samples from brown-headed spider monkeys and mantled howler monkeys. In the laboratory, we dissected and removed the seeds from these dung ball and fecal samples (Ndungball = 3659 seeds and Nfecal = 2793 seeds). The seeds from both the dung balls and fecal samples were then grouped by morphospecies, and the seeds belonging to each morphospecies within each sample were counted. We determined morphospecies visually, and seeds that look similar throughout both the data set of seeds in dung balls and fecal samples were grouped in the same morphospecies. For each morphospecies, we measured seed length (longest side to the nearest millimeter) and seed width (widest point 90 degrees from length) for up to 20 seeds per morphospecies (as is common in studies of this nature, there are many singlets). We also characterized seed surface as pubescent, striate, or smooth. Pubescent seeds are defined by hairs, or hair-like projections coming from their surface, striate seeds by their grooves or indentations in their surface, smooth seeds by their relatively uninterrupted surface. The shape was characterized with a roundness index R(roundness) = L/W where L is the length of seed along the longest axis and W is the widest part of the seed at 90° from L. The closer to 1, the more rounded the seed is, and a larger number correspond to a more elongated shape. Additionally, seed source (fecal sample or dung ball), monkey species, and beetle species were also recorded for each sample. From the set of these seed traits and sources, we examined how dung beetles filter seeds from feces by comparing the traits of seeds in dung balls to the traits of seeds in fecal samples. We also related the range of seed sizes in dung balls of both O. conspicillatum and the smaller C. angustatus. We used a Mann–Whitney U test to compare morphospecies average values in all cases. We then used the seed length as a variable to predict the roundness, using Spearman's rank correlation coefficient. Finally, we calculated the proportion of large pubescent seeds in a dung ball or a fecal sample using the set of seeds in dung balls, compared to seeds in fecal samples as a reference. For both proportions, we calculated 95% confidence intervals. To define large seeds, we used the upper quartile of morphospecies length.
2.2 Seed traits from literatureTo test whether the seed traits for our subset of seeds are similar to the wider Neotropics, we compiled a data base of Neotropical seeds from 123 families and 427 genera, which are not wind dispersed from the reference work “Seeds of Amazonian Plants” (Cornejo & Janovec, 2010). We then added dispersal type data from the Royal Botanic Gardens Kew Seed Information Database (SID, Royal Botanic Gardens Kew, 2020) and our field observations where available. This date set covers a much larger geographic area than our study and could indicate if the results extend beyond our study site. Our final seed trait data base consisted of seed shape, size class, seed surface, and type of seed dispersal. Only mammal dispersed genera or families were included in our statistical analysis (70 families and 176 genera). We eliminated genera without mammal dispersal, or dispersal data, because if they are not in mammal feces, they are most likely not secondarily dispersed by dung beetles. It should be noted here that this might be too conservative because we eliminated over a half of the genera and many trees in tropical forests are mammal dispersed (Muller-Landau & Hardesty, 2005). Seed trait values are categorical and were predetermined by the categories from the “Seeds of Amazonian plants” or the KEW data base. Seed shape was broken into four categories: (1) elongate (1.5 times as long as wide), (2) round (less than 1.5 times as long as wide), (3) flat (width is 1.4 times thickness), and finally (4) irregular (a shape that cannot be easily measured or categorized). Size classes were broken down as follows: 0–0.49, 0.5–0.99, 1–1.99 cm, and finally greater than or equal to 2.00 cm. Seed surface categories for statistical analysis were pubescent vs non-pubescent, similar to the previously described categories but for this analysis we combined the categories smooth and striate.
We tested the following predictions on this data set. (1) Large endozoochorous seeds have a disproportionate number of pubescent genera (or families) because they are unlikely to be incidentally included in dung balls, unlike small seeds (prediction 3 introduction). (2) Rounded seeds should have a disproportionate number of pubescent genera (or families), because of their increased “rollability.” (3) The combination of large size and round shape should be an even better predictor of seed pubescence. To test trait associations, we created contingency tables with the number of genera (or families) of pubescent/non-pubescent seeds in each size or shape class. We then used the Patefield's algorithm to test whether associations were significant, for example, to evaluate whether pubescence occurred more often than predicted by chance in large or rounded seeds (Patefield, 1981). In conjunction with the null model, the H20 value was used as an indicator of the strength of trait partitioning (Blüthgen et al., 2006) (range from 0 to 1, with 0 showing traits are evenly distributed and 1 is the maximal degree of trait partitioning given the totals). Our first and second contingency tables test (1) whether rounded seeds are disproportionately pubescent and (2) whether larger seeds are disproportionately pubescent, respectively. Finally, we created a third contingency table to test whether the combination of roundness and large size better predict pubescence. The latter table was created using genera with the following trait combinations: (1) Seeds larger than 0.99 cm, elongate or round in shape and pubescent, (2) all other pubescent seeds, (3) large seeds, round or elongate but non-pubescent, (4) all other seeds (Table 1).
TABLE 1. Contingency table showing the number of plant genera with particular combinations of seed traits (i.e., large size greater than 0.99 cm, round or elongate) with the presence or absence of pubescence Large round or elongate seeds All other seeds Pubescent seeds 17a 7 Non-pubescent seeds 112 132 a Spondias is in this cell of the contingency table. 3 RESULTS 3.1 Field observations 3.1.1 Natural history observationOn the 20th of March 2019, around 1100 h, during the opportunistic collection of dung beetle brood balls from the dung of brown-headed spider monkeys (A. fusciceps fusciceps) and mantled howler monkeys (A. palliata), we observed three different dung beetle species interacting with large (≈2.1 cm long) seeds from S. mombin as though they were dung. The behavior of three different dung beetles is described here. Each dung beetle handled dung and the seeds differently. First, we observed O. conspicillatum (2.5 cm body length) pushing S. mombin (Figure 1b). Oxysternon conspicillatum is a common tunneling dung beetle in the Reserva Canandé. Second, we observed C. angustatus (0.7 cm body length) trying to pull a section of dung off the S. mombin seed to form a ball. We observed them removing seeds of about 0.5–1 cm length from their balls and then rolling their ball on multiple occasions. Finally, we observed Canthidium sp. (0.6 cm body length) tunnel under the S. mombin (Figure 1a). Occasionally, O. conspicillatum and C. angustatus abandoned the seed after they could not remove dung from the seed. The pubescent (hairy) surface of S. mombin holds dung more effectively than smooth seeds of equivalent size and shape in the same site. During our field observations, dung beetles never accepted non-pubescent, endozoochorous seeds of the same size as S. mombin.
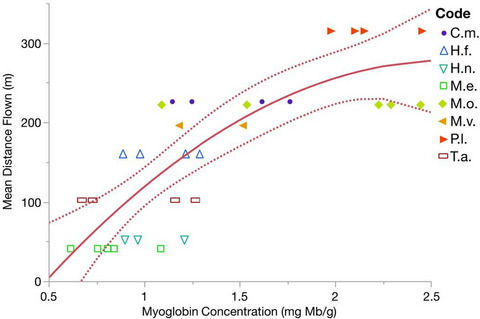
3.2 Morphospecies seed traits 3.2.1 Beetle size and seed lengthOur prediction (1) that larger dung beetles accept larger seeds in their dung balls was confirmed. We first tested whether seed morphospecies in dung balls were smaller than seed morphospecies in the reference fecal set. The mean length of morphospecies in fecal samples (N = 31, M = 6.6 mm ± SD = 6.7 mm) was 1.6 times longer than morphospecies in dung balls (N = 35, M = 4.2 ± 4.8 mm), but this difference was not significant using a Mann–Whitney U test (W = 637.5, df = 1, p = .22). To test whether the larger beetle O. conspicillatum accepts larger morphospecies than the smaller C. angustatus, we compared the mean length of morphospecies in both beetle's dung balls. The length of morphospecies acceptable to O. conspicillatum (N = 14, M = 6.8 ± 6.3 mm) was a statistically significant 2.7 times longer than morphospecies acceptable to C. angustatus (N = 29, M = 2.5 ± 2.1 mm, W = 108, p = .014). However, most seeds in the dung balls of both species were small, and when means were weighted by the number of seeds in each morphospecies, the mean seed lengths from both species were smaller than the unweighted means (M = 4 mm and M = 1.7 mm) for O. conspicillatum and C. angustatus. To test whether the set of morphospecies in each beetle's dung balls differed from morphospecies in the set of fecal samples, we compared the lengths of morphospecies within dung balls of the smaller C. angustatus and the larger O. conspicillatum to the lengths of morphospecies in fecal samples. The mean length of morphospecies in C. angustatus dung balls was 2.6 times shorter than seeds in fecal samples and the difference is statistically significant (W = 303, p = .03). As expected, the range of seed sizes in C. angustatus dung balls (min = 0.7 mm, max = 11 mm) was much smaller than the size range of fecal samples (min = 0.6 mm, max = 23 mm). Oxysternon conspicillatum, in contrast, accepted longer morphospecies, with a mean morphospecies length similar to that of fecal samples (W = 235.5, p = .66). The range of morphospecies lengths was also similar to the range seen in fecal samples (min = 1 mm, max = 20.5 mm) (Figure 2).
 Distribution of seed size across samples. Each plant morphospecies is represented by a data point. Pubescent morphospecies are denoted with an asterisk, smooth morphospecies are denoted by a filled circle, and striate morphospecies are indicated with an inverted triangle. Beetles are placed along the x-axis at a height that indicates their body length. Seed length is plotted on a log scale. The arrow points to an outlier seed in Canthon angustatus dung balls which was flat, and possibly pubescent, though it is unclear as the surface was damaged (Figure S1). Image of seed is provided in Supporting Information.
3.2.2 Morphospecies and shape
Distribution of seed size across samples. Each plant morphospecies is represented by a data point. Pubescent morphospecies are denoted with an asterisk, smooth morphospecies are denoted by a filled circle, and striate morphospecies are indicated with an inverted triangle. Beetles are placed along the x-axis at a height that indicates their body length. Seed length is plotted on a log scale. The arrow points to an outlier seed in Canthon angustatus dung balls which was flat, and possibly pubescent, though it is unclear as the surface was damaged (Figure S1). Image of seed is provided in Supporting Information.
3.2.2 Morphospecies and shape
We tested the prediction (2) that large endozoochorous seeds were rounder and did find a weak correlation between larger seed size and a less elongated shape (ρ = −0.19, p = .15; Figure 4a) though it was not significant. We also tested whether the shape of pubescent seeds was different from non-pubescent seeds. Pubescent morphospecies (N = 17, M = 1.6 ± 0.48) were not significantly different in roundness to non-pubescent morphospecies (N = 38, M = 1.8, SD = 0.60) (W = 280, p = .44; Figure 3b). We also compared the shape of morphospecies in dung balls with the reference to morphospecies in fecal samples. Morphospecies in dung balls (N = 35, M = 1.7 ± 0.59) were not significantly different in shape from morphospecies in fecal samples (N = 31, M = 1.7 ± 0.45) (W = 638, p = .22).

(a) Range of size and surface traits of seeds from fecal samples and dung balls. Pubescent seeds were larger than either smooth or striate seeds. A photograph of a typical seed for each surface type is provided. (b) When we categorized seeds in the same way as in the literature, rounded and elongated seeds were represented by a large number of morphospecies
3.2.3 Morphospecies length and surfaceOur prediction (3) that large endozoochorous seeds species have more pubescent seed species than smaller seed species was supported. We compared the mean length of pubescent morphospecies with non-pubescent morphospecies. Pubescent morphospecies (M = 8.7 ± 6.8 mm) were a statistically significant 2.23 times longer than non-pubescent morphospecies (M = 3.8 mm, SD = 4.5 mm, W = 513, p < .001; Figure 4b). To clarify whether the smaller mean of non-pubescent species was associated with smooth or striate surfaces, we also compared the mean length of pubescent versus smooth and striate morphospecies. Smooth morphospecies (N = 18, M = 3.9 ± 4.1 mm) were 1.5 times shorter than pubescent ones, which is significant (W = 236, p < .01). The mean length of striate morphospecies (M = 22 3.6 ± 4.7 mm) and the mean length of smooth morphospecies (W = 213, p = .68) are not significant. Both are shorter than pubescent morphospecies (W = 304, p < .001; Figure 3a).

(a) Size and roundness were weakly correlated. The length of Canthon angustatus and Oxysternon conspicillatum is indicated on the x-axis. The shaded green area (below 1.5) marks “round” seeds based on the literature definition. (b) Size distributions of non-pubescent morphospecies where smaller seeds make up a larger proportion, whereas (c) pubescent morphospecies showed a clear trend toward more morphospecies with increasing size categories within the data set
Our final prediction (4) that large pubescent seeds (upper quartile) would be over-represented in dung balls in relation to fecal samples was supported. There were 52 large seeds in 21 fecal samples of which 20 were pubescent; hence, the proportion of pubescent seeds in fecal samples was 0.38 95% CI [0.17, 0.59]. The proportion of large pubescent seeds in dung balls was much higher, with 0.94 95% CI [0.80, 1.0], comprising 23 dung balls were made by 11 beetles with 84 large seeds, of which 79 were pubescent and statistically significantly different.
3.3 Seed shape and pubescence from the literatureUsing the trait data from “Seeds of Amazonian Plants” and the dispersal data from KEW SID (Cornejo & Janovec, 2010) to examine patterns at the family and genus level, we found that seeds with a combination of large size and rounded shape at the genus level are slightly more likely to be pubescent than other seeds (H20 = 0.147, p = .048). However, neither shape (H20 = 0.011, p = .26) nor size (H20 = 0.056, p = .13) alone were sufficient to predict pubescence. The pattern was similar at the family level (shape: H20 = 0.014, p = .35, size: H20 = 0.049, p = .13; Figure 5). However, if we do not exclude all genera without dispersal data from our data set, both size and the combination of size and shape are significant predictors of seed pubescence at the genus level (Tables S1–S3).

Neotropical seeds with zoochorous mammal dispersal, and the distribution of genera belonging to each shape and size class. Pubescent seeds in each size or shape class are in black, and all other seeds are in gray. (a) The number of genera in each shape category. Shape was not strongly associated with seed pubescence H20 = 0.011, p = .26. (b) The number of genera in each size class. Seed size was also not coupled to seed pubescence, although trait syndromes showed a higher degree of association than for shape H20 = 0.056, p = .12
4 DISCUSSIONEndozoochory is the dominate form of seed dispersal in the Neotropics, with 65%−100% of seeds being dispersed by animals (Muller-Landau & Hardesty, 2005; Seidler & Plotkin, 2006). Primates are important seed dispersers, and seeds passing through their digestive tract like seeds in our study have an equal or greater chance of germinating (Fuzessy et al., 2016; Stevenson et al., 2002), suggesting that this is a desirable trait for of seed dispersal. This may be augmented by secondary dispersal by dung beetles because of the aggregated deposition of seeds by some primates (e.g., Russo & Augspurger, 2004). At least one study of tamarins and associated dung beetles did find some increase in rates of germination with secondary dispersal by dung beetles in secondary forests (Culot et al., 2018) and in forests like our study site where spider monkeys are one of the most important frugivores secondary dispersal by dung beetles might be even more important because of the high density of seeds in their dung.
We examined seed traits in relation to dung beetle acceptance of seeds in their dung balls. Our results confirmed the importance of seed size which has been the main trait considered previously (Andresen, 2002; Andresen & Levey, 2004; Braga et al., 2017). In addition, we also highlighted two traits (seed surface and seed shape) that have not previously been studied concerning secondary dispersal. We considered both in situ observations of seed inclusion in dung balls and overall trends in seed trait associations from the literature. Our results support our hypothesis that seed traits size and pubescence facilitate secondary seed dispersal by dung beetles, but not shape. Three out of four predictions from our hypothesis were supported.
Our data are also consistent with the prediction (1) that larger beetles accept larger seeds (Andresen, 2001). Oxysternon conspicillatum is ≈3.6 times longer than C. angustatus, and the average length of morphospecies in O. conspicillatum dung balls was almost three times the mean length of seeds in C. angustatus dung balls. The maximum acceptable seed length for both beetles was close to their body length. Together, both beetles covered the entire range of seed sizes found in the feces of both howler and spider monkeys (Figure 2). This has ecological consequences for those seeds.
One of the benefits of secondary dispersal is escape from seed predation, and burial even at shallow depths of 1 cm has been shown to drastically reduce seed removal by rodents (Feer, 1999). The greater the burial depth, the lower the chance that a seed is removed by a seed predator, but the chance that a seed will germinate may be reduced. Small seeds are both less likely to germinate and more likely to be buried at greater depths which forms scenario where secondary dispersal may be detrimental. Small seeds are much less likely to germinate at depths greater than 3 cm, and large seeds are unlikely to germinate at depths greater than 5 cm (Shepherd & Chapman, 1998). Previously recorded seed burial depths by a Canthon species equal 4 cm for seeds smaller than 5 mm. They did not bury larger seeds. For O. conspicillatum, mean burial depths for seeds <5 mm were 5.65 cm and for larger seeds 2.57 cm (Vulinec, 2002). However, the reduced density of seeds on the surface or at lower depths might still create a net benefit for hundreds of small seeds (<2 mm) found in some of our spider monkey fecal samples.
Our prediction (2) that large mammal dispersed seeds are rounded was not supported and most seeds, regardless of size, tended to be somewhat rounded (Figure 4a). Additionally, seeds in dung balls did not differ in shape compared to seeds in fecal samples, suggesting that this may not be the most crucial seed trait for inclusion in dung balls by dung beetles. While shape was not found to be important here, it is possible that the number of morphospecies we have in our data set is too small because there does appear to be a trend toward less elongate seeds at larger sizes. In addition, it was an important variable in the literature data though only in combination with shape and size.
Our prediction (3) that large seeds are more pubescent than smaller seeds was more strongly supported by the field data (Figure 4b). At the genus level (literature data), the combination of large size and pubescence occurred more often than expected by chance suggesting that the trend might extend beyond our study system. However, further work is required. We also excluded many genera from the literature data set for the lack of dispersal data. This may be a mistake given that the majority of Neotropical trees are animal dispersed. A more complete data set with a continuous size variable and more complete dispersal data is needed. Finally, our prediction (4) that large pubescent seeds are more acceptable to dung beetles than large non-pubescent seeds was supported by our direct field observations of O. conspicillatum, C. angustatus, and Canthidium sp. and their interaction with S. mombin. In addition, large seeds in dung balls were disproportionately pubescent, especially when compared to the reference set of seeds in fecal samples. The disproportionate acceptance of large pubescent seeds by dung beetles indicates that they filter seeds with respect to both size and surface. Together, these findings suggest that this combination of seed traits is advantageous for secondary dispersal.
Of the 11 species of dung beetles which we observed on primate dung over 6 months, three were observed to treat the dung covered S. mombin as though it were feces. This unproductive foraging likely led to a delay in a feeding or reproductive opportunity, while the surrounding dung was buried by competitors (Figure 1). The cost may not be severe, because monkeys are a reliable source of dung, and we often saw dung beetles perch on leaves below monkey troops, apparently waiting for them to defecate. Despite S. mombin being 1.6 times longer than the next largest acceptable seed included in O. conspicillatum dung balls, and 2.3 times than even the longest outlier in C. angustatus balls, both beetles were attracted to and attempted to move S. mombin seeds. The combination of seed traits, large size, roundness, and pubescence, appears to make S. mombin an ideal candidate for secondary dispersal by large dung beetles.
Large seeds representing a larger parental investment (K-selection rather than r-selection) are more likely to respond strongly to conditions that increase survival. Secondary dispersal and burial by dung beetles increase their distance from conspecifics and their protection against seed predators and pathogens. These benefits could favor pubescence and thus dung attachment to seeds. Most mammals consume multiple fruits from the same tree. Personal observations in the field suggest there is a lot of synchronous defecation of howler and spider monkeys. Consequently, seeds from the same tree are often concentrated together within the mammal dung which should increase the relative benefits of secondary dispersal (Howe & Smallwood, 1982; Seidler & Plotkin, 2006). In the case of S. mombin, we observed many seeds falling close to each other, suggesting that dispersal away from conspecifics should be beneficial. However, in the case of small primates (e.g., tamarins), seeds may be deposited singly or in small numbers (Culot et al., 2018), which would weaken selective pressure for secondary dispersal by dung beetles because there is no dung to attract the dung beetles. In cases where there is dung it acts like a glue sticking seeds together. In these cases, dung beetles should be the most beneficial. For this reason, further study is required to fully understand in which habitats, and with which monkey species dung beetles are most beneficial as secondary dispersers.
Our evidence from the literature data that larger seeds are more likely to be pubescent suggests that this trend extends to other sites in the Neotropics, which will require further local studies. At least for primate-dispersed seeds, however, we would expect a weaker trend in the old world because of a larger number of seed spitting primates (Dominy & Duncan, 2005). When seeds are spat, they do not pass through the digestive system of a primate and are thus not in the dung for dung beetles to interact with. Mammals are more likely to spit larger seeds; hence, seed spitting may blur the relative advantage of pubescence.
There are examples of dung beetle dispersal syndrome. One example is of C. argenteum seeds rolled by the Epirinus flagellatus dung beetles in De Hoop Nature Reserve, South Africa. Seeds with morphologies similar to C. argenteum are exposed on the surface and dispersed or eaten by small rodents. Instead, C. argenteum seeds smell strongly of herbivore feces and are dispersed and buried by dung beetles (Midgley et al., 2015). For S. mombin and other pubescent seeds, we suggest that these seeds use the actual dung to increase their attractiveness to dung beetles. Another example comes from an observation in the Ivory Coast where 26 dung beetles from five species mistook single seeds as dung, though in the observation seeds were only characterized by size (Kunz & Krell, 2011). Our results together with the aforementioned studies suggest that this phenomenon deserves further study.
5 CONCLUSIONDung beetles treat the S. mombin seeds similarly to dung, attempt to move the whole seed, or remove a part of it to form a ball, even though the seed is well outside the typical, acceptable seed size range for the beetle species. We suggest some cost to the beetle because of the lost opportunity and the need to spend more time searching for new dung. The finding that large pubescent seeds were accepted while large smooth seeds were excluded suggests that surfaces are essential. It is still unknown how widespread this phenomenon is. The combination of large size and a rounder, easier to roll shape corresponds to an increased proportion of pubescence across plant genera. Also, pubescence and shape may be variable traits in some seeds, capable of responding to environmental conditions. A clumped distribution and predation risk may be particularly costly to large seeds where secondary dispersal by dung beetles is particularly beneficial. We suggest that additional observations with a particular focus on large, round, pubescent seeds in the genera listed in Table 2 would be useful to elucidate how important this phenomenon is in the evolution of seed traits.
TABLE 2. Genera of large, rounded, pubescent seeds that might exploit dung beetles as secondary dispersers in a similar manner as Spondias mombin Family Genus Size class Shape Endozoochory Anacardiaceae Spondias 2+ Round Yes Arecaceae Astrocaryum 2+ Round Yes Arecaceae Attalea 2+ Elongate Yes Arecaceae Bactris 0.5–2+ Elongate Yes Arecaceae Chamaedorea 1.00–1.99 Elongate Yes Arecaceae Desmoncus 1.00–1.99 Elongate Yes Arecaceae Euterpe 1.00–1.99 Round Yes Arecaceae Oenocarpus 1.00–2+ Round Yes Bombacaceae Matisia 2+ Elongate Yes Bombacaceae Quararibea 1.00–1.99 Elongate Yes Caryocaraceae Caryocar 2+ Round Yes Chrysobalanaceae Couepia 2+ Elongate Yes Dipteryx Dipteryx 2+ Elongate Yes Quiinaceae Quiina 1.00–1.99 Elongate Yes Verbenaceae Vitex 1.00–1.99 Elongate Yes Violaceae Rinorea 0.5–0.99 Round Yes ACKNOWLEDGMENTWe would like to thank the Jocotoco, the Reserva Canandé, and Tesoro Escondido for assistance in the field, Andrea Hilpert for dissecting so many dung balls and sorting seeds, Juan Ernesto Guevara for seed identification, Eckhard Heymann for helpful comments and suggestions, and Maxim Ionov for his comments and assistance in the preparation of this manuscript. In addition, we would like to thank Citlalli Morelos-Juarez for her referance list of plants spider monkeys feed on. Finally for help in the field we would like to thank, Adriana Argoti Avila, Bryan Xavier Tamayo Zambrano, Jorge Alipio Zambrano Velez, José Amado De la Cruz Chávez, José Roberto de la Cruz Loor, Alcides Agustín Zambrano Velez, José Manuel Añapa Añapa, Ronaldo Mesías, Vanessa Moreira, Daniel Velázquez, and Yadira Giler. Open Access funding enabled and organized by Projekt DEAL.
留言 (0)