
記住我
Actin is one of the most abundant proteins in eukaryotic cells and owing to its ability to polymerise into filaments (F-Actin) forming static or highly dynamic networks, plays an important function in many crucial cellular processes. The regulated dynamics of F-actin and crosslinking of individual filaments requires the integration of actin binding proteins (ABPs) with signalling cascades that ultimately regulate actin filament length, stability and anchorage (Hansen & Kwiatkowski, 2013). This control can be achieved at multiple levels, from direct binding of ABPs to post-translational modifications of actin and are based on influencing the basic polymerisation mechanism of this molecule (Dominguez & Holmes, 2011).
The atomic structure of monomeric G-actin was first described in 1990. With a molecular weight of 42 kDa (Pollard, 2016), the monomer consists of four subdomains (Kabsch, Mannherz, Suck, Pai, & Holmes, 1990); G-actin binds ATP or ADP in the cleft between subdomains 2 and 4 (Graceffa & Dominguez, 2003; Kabsch et al., 1990; Otterbein, Graceffa, & Dominguez, 2001). G-actin-ATP and G-actin-ADP show conformational differences in subdomain 2 (Otterbein et al., 2001; Graceffa & Dominguez, 2003), imparting G-actin-ATP the ability to bind other ATP-bound actin monomers, facilitating polymerisation into F-actin (Pollard, 1984). Studies using electron microscopy in 1963 proposed that F-actin consists of two actin strands that intertwine to form a right-handed helical filament (Hanson, 1963). More recently, the F-actin structure was described by cryo-electron microscopy (Fujii, Iwane, Yanagida, & Namba, 2010; von der Ecken, Heissler, Pathan-Chhatbar, Manstein, & Raunser, 2016) and it has been suggested that it represents a single left-handed helix (Dominguez & Holmes, 2011). Dominguez and Holmes argued that, since the twist per actin molecule is −166° (close to −180°), the F-actin filament structure only appears two-stranded that slowly turns in a right-handed fashion. Hence, the F-actin structure can either be described as a two-start right-handed helix or a single-start left-handed helix with a diameter close to 7–10 nm (Dominguez & Holmes, 2011). These arrangements of F-actin can result into different configurations and includes filaments orientated in the same direction, parallel and antiparallel bundles, and filaments forming actin lattices that enable it to fulfil different mechanical and functional roles (Chhabra & Higgs, 2007).
Actin polymerisation is a dynamic process that involves rapid changes in the assembly and disassembly of F-actin. One critical factor for the transition between G-actin and F-actin (actin treadmilling) is the hydrolysis of ATP (Korn, Carlier, & Pantaloni, 1987). The actin treadmilling cycle starts with the addition of G-actin-ATP to “barbed” (plus) end of the F-actin filament. During this polymerisation step, ATP is hydrolysed to ADP and inorganic phosphate (Pi). Hydrolysis of ATP results in stable F-actin filaments with bound ADP + Pi (F-actin-ADP-Pi). Slow release of Pi results in F-actin-ADP and destabilises the filament, resulting in the release of G-actin-ADP from the filament. In a growing F-actin strand, F-actin-ADP resides at the “pointed” (minus) end where the depolymerisation of G-actin-ADP occurs. Subsequently, ADP can be replaced with ATP in the monomer, generating a new G-actin-ATP molecule. The addition of a new G-actin-ATP molecule to the F-actin barbed end causes a conformational change in the adjacent actin molecule. Murakami and co-workers proposed this structural change to be responsible for the initiation of ATP hydrolysis (Murakami et al., 2010). Within subdomain 2 of the actin molecule lies the DNase I binding (D-) loop, which inserts itself into the hydrophobic cleft between subdomains 1 and 3 of the apposing monomer and is crucial for polymerisation (Fujii et al., 2010). Important differences in the amino acid residues of the D-loop between canonical and apicomplexan actins give rise to different filament polymerisation dynamics, which will be discussed in Section 2.2.
Of relevance is that the amount of G-actin has to be above a certain concentration threshold for polymerisation to occur. This threshold is referred to as the critical concentration, where the removal of G-actin occurs at the same rate as the addition of new monomers, but the net mass of the F-actin polymer does not change (Pollard & Borisy, 2003). Importantly, the critical concentration for polymerisation to happen at the barbed end is lower than the concentration needed for polymerisation at the pointed end or for the formation of an actin dimer (Pollard & Borisy, 2003; Pollard, 2016). This results in the typical behaviour of F-actin, where monomers at the barbed end are added 5–10 times faster than to the slow-growing pointed end. Together, this cooperative polymerisation process results in the so called treadmillling of actin filaments (Figure 1a). It should also be mentioned that the critical concentration required for G-actin-ATP to polymerise was reported to be lower than the concentration required for G-actin-ADP (Cooke, 1975; Pollard, 1984). Consequently, actin polymerisation is more likely to occur at the barbed end of an F-actin strand by attaching G-actin-ATP monomers.

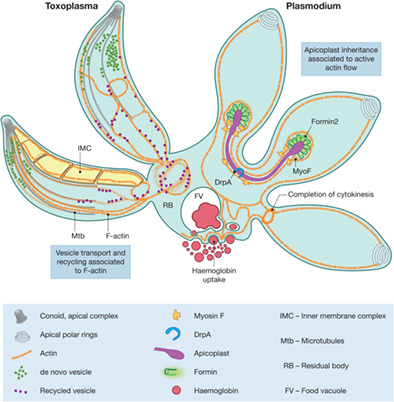
Host cell invasion and intracellular F-actin flow. (a) F-actin polymer growth depends on a critical concentration of G-actin in the cytoplasm. The balance between incorporation of monomers during elongation and depolymerisation is controlled by multiple regulators. At the barbed end, profilin delivers ATP-rich monomers to formins for nucleation and polymer elongation; at the pointed end, ADF-1 binds ADP-rich actin and contributes to depolymerisation, polymer fragmentation and ATP hydrolysis. Nucleotide exchange on released ADP-actin to form ATP-actin for a new round of polymerisation could be brought about by CAP and profilin. Stability of F-actin bundles is maintained by several actin binding proteins including CPs, coronins and formins. (b) The linear model for parasite motility proposes that a motor complex consisting of actin, myosins and associated proteins is located within the narrow space of 20 nm between the plasma membrane and the underneath saccule-like structure, the IMC. (c) Other alternative models can be proposed in which cytosolic myosins and IMC permeability contribute to transport of microneme adhesins and exchange of actin between the cytosol and space underneath the plasma membrane. (d) Cytoplasmic Actin and cell invasion. Initial attachment to the surface of a host cell depends on F-actin accumulation at the posterior pole and at the apical end. During invasion, the tight junction complex (TJ) is formed that stabilises the attachment of the parasite to the host cell. F-actin at the TJ is involved in force generation and parasite stability to facilitate nuclear entry. F-actin at the posterior end produces an actin lattice that provides plasticity and a “pushing” contraction force to allow nuclear deformation and entry through the narrow space of the tight junction. Deformation and contraction of the parasite region lacking the rigid structure of microtubules might facilitate nuclear entry by “pulling.” We propose that the nucleus is squeezed through the TJ by a push–pull mechanism controlled by actin and potentially microtubules. (e) F-actin flow model in the parasitophorous vacuole during endodyogeny. There are three formins identified that have specific localisations: Formin-1 is localized at the apical end, formin-2 is closely associated to apicoplasts and formin-3 is at the parasite posterior end. Flow analysis combined with formin conditional knock-out studies suggest that F-actin flow is controlled by the formins. Formin-1 controls the apical posterior flow direction, formin-2 may be responsible for bidirectional flow of actin, and formin-3 may be responsible for controlling the flow in the RB and retrograde flow in the parasite. (f) Actin flow is involved in apicoplast inheritance in Apicomplexa. In Plasmodium, the actin network additionally regulates the transport of endocytosed haemoglobin to the food vacuole/fusion of vesicles, and the completion of cytokinesis
The first step of de novo F-actin filament formation is called actin nucleation, where a new F-actin filament is assembled from G-actin monomers (Pollard, Blanchoin, & Mullins, 2000). Inevitably, this process requires the formation of actin dimers and trimers, a process that is kinetically unfavourable (Pollard, 2016; Sept & McCammon, 2001). From this point onwards, G-actin monomer assembly to the actin trimer occurs with the same rate as G-actin polymerisation on existing F-actin filaments (Sept & McCammon, 2001). The assembly of this polymerisation nucleus therefore represents the critical step that has to be overcome for de novo F-actin formation (Sept & McCammon, 2001). To efficiently control kinetically unfavourable steps of actin dynamics, a vast number of actin binding proteins (ABPs) has been described, some highly conserved in most eukaryotes and others unique to certain species (see [Pollard, 2016] and Table 1). In contrast to the huge repertoire of ABPs found in higher eukaryotes, only a handful of ABPs have been identified in most protozoa, including apicomplexan parasites (Baum, Papenfuss, Baum, Speed, & Cowman, 2006; Sattler et al., 2011; Schuler & Matuschewski, 2006) (Table 1).
TABLE 1. Summary of main roles of ABP and ABP regulatory protein partners Higher eukaryotes Apicomplexa Function Protein ABP regulatory proteins Polymer Actin isoform P. falciparum T.gondii C. parvumα-skeletal
α-smooth
α-cardiac
β-cytoplasmic
γ-enteric smooth muscle
γ-cytoplasmic
Pf actin 1
Pf actin 2
Tg actin Cp actin Monomer binding ProfilinPf profilin
Pf CAP-like
Tg profilin
Tg CAP-like
Cp profilin
Cp CAP-like
Thymosin-β4 WH2 (WASP-homology 2) Polymerase Formin family proteins Rho GTPasesPf Form1
Pf Form2
Tg Form1
Tg Form2
Tg Form3
Cp Form1
Cp Form2
ENA VASP Nucleation Initiation Formin isoforms (~15) Branching Arp2/3WASH/WASP
Reg complex;
DIP1-Arpin
WASp, N-WASPPolyphosphoinositoides,
SRC homology 3 domains
Rho- GTPases,
SCAR/WAVE SCAR/WAVE reg complex and RAC GTPases Lmod Alternative filament formationTandem
WH-2
(spire
Cordon bleu, JMY)
Cofilin
CP loss
Pf actin-like PfMTRAP-CT
PfRH1-CT
Tg actin-like Cp actin-like Capping proteins Barbed ends CPCARMIL
Phosphoinositoides
Pf CAPα
Pf CAPβ
Tg CAP α
Tg CAP β
Cp CAPα
Cp CAPβ
Pointed endsTropomodulin
Tropomyosin
Arp2/3
Severing proteinsCofilin 1
Muscle-specific MS cofilin2
Gelsolin family (barbed end)
Formin
FRL-α
Formin
INF-2
AIP1
Pf ADF1
Pf ADF2
Pf Chronophin
TgADF1
Tg Chronophin
Cp ADF1 Pf14-3-3 Pf14-3-3 Cp14-3-3 Cross-linkingFimbrin
Filamin
Pf Coronin Tg Coronin Cp Coronin Calponin-domains Note: Adapted from Schuler and Matuschewski (2006), Pollard (2016), Sattler, Ganter, Hliscs, Matuschewski, and Schuler (2011). Abbreviations: ALP, actin like protein; CAP like C-terminal extended-sheet domain of cyclase associated proteins; CAP, capping protein; Cp, Cryptosporidium parvum; Form, formin; Pf, Plasmodium falciparum; Tg, Toxoplasma gondii.The formation of actin polymers is a tightly regulated process that involves several steps: Nucleation, polymerisation and regulation of the polymer size by actin treadmilling and filament stabilisation. Nucleation of canonical actins depends on a set of ABPs to overcome the activation energy barrier of de novo filament formation. Three different types of actin nucleators have been described: the Arp2/3 complex, spire and the formin protein family (Goode & Eck, 2007). The Arp2/3 complex consists of seven subunits and promotes branching and formation of novel daughter F-actin filaments at an angle of 70° from an already existing filament (Mullins, Heuser, & Pollard, 1998). The Arp2/3 complex is an important nucleator of F-actin in lamellipodia and is involved in cell migration (Suraneni et al., 2012). Spire, first discovered in Drosophila, possesses four WH2 domains used for attracting four G-actin monomers to create a nucleation complex, and can collaborate with formin to build essential cytoskeletal structures (Quinlan, Heuser, Kerkhoff, & Mullins, 2005). Formins contain two formin homology (FH) domains, FH1 and FH2. The FH2 domain nucleates and elongates unbranched actin filaments by “processive capping” at the barbed end, while the FH1 domain can interact with the ABP profilin, release actin monomers sequested by profilin and incorporate them into the growing filament (Courtemanche, 2018). In apicomplexans, only formin like proteins have been identified and are thought to represent the only actin nucleators in these parasites (Tosetti, Dos Santos Pacheco, Soldati-Favre, & Jacot, 2019).
Actin treadmilling is the continuous removal of monomers from the pointed ends of filaments and their simultaneous incorporation at the barbed end, a process in which the ABPs formin, actin depolymerizing factor (ADF)/cofilin and profilin are involved. ADF/cofilin family bind and destabilise F-actin filaments, thus increasing the amount of available G-actin monomers (Moon & Drubin, 1995; Nishida, Maekawa, & Sakai, 1984; Yonezawa, Nishida, & Sakai, 1985; Lappalainen & Drubin, 1997). Cofilin and ADF have higher affinity to G-Actin-ADP than to G-Actin-ATP, thereby increasing the depolymerisation rate of F-actin. While depolymerisation occurs at the pointed end during the treadmilling process, F-actin filament elongation takes place at the barbed end. One of the proteins involved in mediating filament assembly is the polymerisation factor profilin, which binds monomeric G-actin (Baum et al., 2006; Pollard, 2016; Carlsson, Nystrom, Sundkvist, Markey, & Lindberg, 1977; Pantaloni & Carlier, 1993). Profilin binds to G-actin-ATP and G-actin-ADP with similar affinity while drastically increasing the exchange rate of actin-bound ADP for ATP (Selden, Kinosian, Estes, & Gershman, 1999). Actin treadmilling occurs by formins assembling a pool of regenerated G-actin-ATP above the critical concentration at the barbed end of an actin polymer (Romero et al., 2004; Kovar, 2006). Cyclase associated protein (CAP) is essential for most eukaryotes and can work in synergy with ADF/cofilin to increase F-actin depolymerization by almost 100-fold, and furthermore, can exchange ADP on depolymerized monomers with ATP to enable another round of F-actin assembly (Kotila et al., 2019). Coronins have been described as a “double-edged sword,” promoting F-actin disassembly in coordination with ADF/cofilin at ADP-rich pointed ends of networks, while promoting rapid F-actin growth at the ATP-rich barbed ends by recruiting the Arp2/3 complex for expansion of branches, thereby functioning during rapid actin-mediated processes auch as endocytosis and cell migration (Gandhi & Goode, 2008). Capping proteins (CPs) are a heretodimer composed of the α and β subunits, bind to barbed ends of F-actin and prevent the addition or removal of monomers, thereby stabilising the filament (Edwards et al., 2014; Pollard, 2016). Interestingly, they also stabilise short filaments produced by the actin related protein 1 (ARP1) (Cooper & Sept, 2008).
2 ACTIN IN APICOMPLEXAN PARASITES 2.1 Introduction to ApicomplexaThe phylum Apicomplexa includes obligate intracellular protozoan parasites such as Plasmodium spp, Toxoplasma gondii and Cyptosporidium spp which cause devastating human and livestock diseases. Actin isoforms have been identified in all apicomplexans; in all of them, the amino acid sequence is highly divergent from canonical actin found in higher eukaryotes with only about 80% amino acid sequence similarity (Baum et al., 2006; Dobrowolski, Niesman, & Sibley, 1997). There are notable differences in apicomplexan actin, in both the number of isoforms and conservation in amino acid sequence. While Toxoplasma (and most other apicomplexans) possesses only a single gene for actin (Dobrowolski et al., 1997), Plasmodium species possess two isoforms, act-1 and act-2 (Wesseling, Smits, & Schoenmakers, 1988). Plasmodium falciparum ACT1 (PfACT1) is expressed in all life cycle stages, while ACT2 was found to be solely expressed in the sexual stages (Wesseling et al., 1989). Toxoplasma gondii actin (TgACT1) shares 93% amino acid sequence identity with PfACT1.
2.2 Dynamics of apicomplexan Actin in vitroWhile in most eukaryotes F-actin can form long filaments in vitro, apicomplexan actin forms only short filaments of less then 100 nm in the absence of filament-stabilising drugs such as jasplakinolide (Pospich et al., 2017; Schmitz et al., 2005). Apicomplexan actin has also been notoriously hard to visualise and characterize, both in vitro and in vivo, leading to conflicting interpretations regarding polymerisation mechanisms and functions. Early studies by Dobrowolski and colleagues used ultra centrifugation methods to investigate the state of the actin polymer (Dobrowolski et al., 1997); the findings failed to detect filaments, leading the authors to propose that actin is mainly in the monomeric G-actin state in Toxoplasma. A comparative study with recombinant actin in vitro showed that Toxoplasma actin formed short unstable filaments (Sahoo, Beatty, Heuser, Sept, & Sibley, 2006). Intriguingly, the critical concentration required for actin polymerisation in Toxoplasma was suggested to be lower compared to conventional actins, while F-actin assembly and turnover was suggested to occur very rapidly. It was proposed that amino acid residues on the Toxoplasma actin monomer surface differ from conventional actins and these differences contribute to filament instability, which could be an adaptation that enables fast parasite motility (Sahoo et al., 2006; Skillman et al., 2011). These results were supported by findings in Plasmodium falciparum (Schmitz et al., 2005), where short filaments (~100 nm) were observed in vitro when compared to rabbit actin (~350 nm). Further experiments using actin sedimentation assays led Skillman and co-workers to speculate on an isodesmic model for polymerisation in apicomplexan parasites, which would be a unique and surprising mechanism, only found in apicomplexans (Skillman et al., 2013). This model predicts that all polymerisation steps occur with the same rate, meaning that no critical concentration exists. In consequence, assembly and disassembly occur at the same rate, including the nucleation step prior to filament elongation. While this might explain the formation of short and highly dynamic actin filaments, it would complicate the regulation of actin dynamics in the cell, making nucleators such as formins redundant.
A recent study demonstrated that sedimentation assays are not reliable for the determination of critical concentration of apicomplexan actins (Kumpula et al., 2017). The isodesmic model has thus been readdressed using alternative technologies. Pyrene fluorescence assays convincingly demonstrated that PfACT1 polymerisation depends on a critical concentration, similar to canonical actins (Kumpula et al., 2017). However, the depolymerisation rate of PfACT1 appears to be very rapid and structural differences compared to canonical actins were proposed to cause instability and fragmentation of F-actin. In agreement, cryo-electron microscopy experiments revealed that the differences in contact sites between PfACT1 molecules were responsible for filament instability (Pospich et al., 2017). Further crystallography studies of PfACT1 identified the Arg178/Asp180-containing A-loop to be one of the factors responsible, which acts as a switch governing the relative stability of F-actin (Kumpula, Lopez, Tajedin, Han, & Kursula, 2019). Finally, a recent study used actin chromobodies and TIRF microscopy to visualise dynamics of PfACT1 (Lu, Fagnant, & Trybus, 2019) and a significantly higher critical concentration was determined. Here, the instability of F-actin was attributed to rapid filament shrinkage at the pointed end (Lu et al., 2019). The PfACT1 D-loop has important differences in amino acid residues compared to canonical actins, which contributes to natural filament instability essential for the parasite (Lu et al., 2019). A chimeric P. berghei actin-1 with a “canonical” D-loop could produce long filaments in vitro and restore gametocytogenesis in parasites lacking actin-2, indicating that the differential functional needs of the two actins rely heavily on differential filament stability (Vahokoski et al., 2014). In another study, a single point mutation N41H within the PfACT1 D-loop allowed PfACT1 incorporation into mammalian F-actin in a skin cell line (Douglas et al., 2018). Taken together, data obtained from recent in vitro experiments demonstrate that apicomplexan F-actin is more unstable than canonical actins. Differences are found in key residues in multiple regions of the G-actin monomers that appear to critically contribute to this phenomenon. Nonetheless, while different critical concentrations have been determined, a cooperative nucleation-elongation mechanism seems to be in place for F-actin polymerisation in apicomplexan parasites, as seen in all other eukaryotes studied thus far.
Interestingly, Theileria annulata parasites possess an actin isoform that has retained the amino acid residues Ser200, Met270 normally seen in canonical actins, which are mutated to Gly200, Lys270 in Plasmodium and Toxoplasma. Perhaps as a consequence, Theileria parasites make more stable F-actin structures, which were detected by cryoelectron tomography (Kuhni-Boghenbor et al., 2012). Indeed, TgACT1 produced more stable filaments with the reverse mutations Gly200Ser and Lys270Met (Skillman et al., 2011).
2.3 Apicomplexan Actin function and distribution in vivoUntil recently it was believed that actin is required primarily for parasite motility. This assumption was based on early inhibitor studies which suggested that microtubules, but not actin, are required for parasite replication (Shaw, He, Roos, & Tilney, 2000) and conversely, actin is required for motility and host cell invasion (Dobrowolski & Sibley, 1996). In the case of Plasmodium, a potential role for PfACT1 in haemoglobin uptake was suggested using inhibitors of actin dynamics (Smythe, Joiner, & Hoppe, 2008). In this study it was concluded that actin has a role in the fusion of endocytic vesicles or in vesicular transport. In Theileria long filamentous nanotubular protrusions of F-actin from the plasma membrane into the host cell cytoplasm were observed, which the authors hypothesise to aid in correct positioning of the parasite within the host cell (Kuhni-Boghenbor et al., 2012).
Multiple, additional functions of F-actin during intracellular parasite development have recently begun to unfold. Using a conditional mutagenesis approach based on DiCre-recombinase it was demonstrated that TgACT1 is essential for maintenance of the apicoplast, an essential apicomplexa-specific chloroplast-like endosymbiotic organelle (Andenmatten et al., 2013; Whitelaw et al., 2017) and maturation of the parasite (Whitelaw et al., 2017). Furthermore, depletion of the unconventional myosin, MyoF, resulted in a similar phenotype, with parasites losing the apicoplast (Heaslip, Nelson, & Warshaw, 2016; Jacot, Daher, & Soldati-Favre, 2013). Importantly, the role of parasite F-actin in apicoplast inheritance appears to be conserved within the phylum, since disruption of pfact1 also resulted in loss of the apicoplast in P. falciparum (Das, Lemgruber, Tay, Baum, & Meissner, 2017).
While these studies suggested that parasite actin is involved in critical intracellular functions, the mechanisms involved remained obscure due to the inability of visualising F-actin dynamics in the parasite. Conventional actin probes, such as phalloidin, GFP-tagged actin, LifeAct or SiR-Act did not allow reliable detection of apicomplexan F-actin, either due to toxicity of the probe or their inability to stain apicomplexan F-actin. Recently, a new approach for the imaging of F-actin based on the expression of actin-chromobodies (Cb) was adapted to apicomlexan parasites. These single chain anti-actin camel antibodies (Rocchetti, Hawes, & Kriechbaumer, 2014) show significantly fewer effects on F-actin dynamics compared to other actin probes in all eukaryotes tested to date (Melak, Plessner, & Grosse, 2017; Panza, Maier, Schmees, Rothbauer, & Sollner, 2015). Using this tool it was demonstrated that F-actin is polymerised by formins in T.gondii and P.falciparum (Stortz et al., 2019; Tosetti et al., 2019) and thereby an extensive intravacuolar network is established that appears to be required for material exchange and cell-to-cell communication between individual parasites. This network is organised by short, bundled F-actin and not by long stable filaments, in good agreement with in vitro studies of apicomplexan actin (Kumpula et al., 2017; Kumpula & Kursula, 2015; Skillman et al., 2011; Skillman et al., 2013). Furthermore, actin nanobodies were tagged with standard fluorescent proteins, photoconvertible proteins mEos3.2 or multifunctional reporters HALO and SNAP enabling their use in diffraction limited microscopy and quantitative super-resolution microscopy methods - dual colour SMLM and live SIM (Periz et al., 2019). In combination with conditional mutants this approach has allowed investigation into the multiple functions of apicomplexan F-actin during the asexual life cycle.
2.4 The role of F-Actin during gliding motility and invasion of the host cellApicomplexan gliding motility and host cell invasion was believed to be a purely parasite actin-driven process (Dobrowolski & Sibley, 1996). According to the linear motor model (Figure 1b), short F-actin filaments are polymerised between the plasma membrane (PM) and the inner membrane complex (IMC, a specialised structure found in apicomplexan parasites that consists of membranous cisternae and structural components located 20–30 nm beneath the PM). These short filaments interact with transmembrane proteins derived from secretion of micronemes (invasion related apical organelles) via the glideosome associated connector (GAC) and the myosin A motor complex that is anchored within the IMC (Jacot et al., 2016). Furthermore, it is believed that during gliding motility and invasion, F-actin is formed at the apical tip of the parasite, where formin-1 is localized (Baum et al., 2008; Jacot et al., 2016). While this model has been supported by several lines of evidence, it cannot explain recent findings that used reverse genetic and biophysical approaches to determine force production and transmission during gliding motility and invasion:
Motility and invasiveness of conditional mutants for core components of the gliding and invasion machinery are affected, but not completely abrogated (Andenmatten et al., 2013; Egarter et al., 2014; Whitelaw et al., 2017) Plasmodium sporozoites do not move in a smooth fashion (as predicted by a linear motor), but in a slip–stick-mechanism, where F-actin appears to be required for the formation and release of attachment sites (Münter et al., 2009) Retrograde membrane flow, a prerequisite for gliding motility, occurs independent of the acto-myosin-system of the parasite (Gras et al., 2019; Quadt, Streichfuss, Moreau, Spatz, & Frischknecht, 2016; Whitelaw et al., 2017). During invasion by Toxoplasma, host cell actin is critical and in case of parasite mutants devoid of the glideosome, a macropinocytosis-like process allows the parasite to invade (Bichet et al., 2016).While one explanation for these observations could be attributed to functional redundancies of paralogs, in particular in the repertoire of myosins and micronemal proteins (Frenal, Marq, Jacot, Polonais, & Soldati-Favre, 2014), the disruption of core components of this system -including the single copy gene for actin- mainly causes defects in the formation and regulation of attachment sites. The effect of these disruptions on gliding and invasion speed are to a much lesser extent (Whitelaw et al., 2017), raising the question whether the acto-myosin system is required as a molecular clutch in order to initiate motility and to transmit the force generated by retrograde membrane flow or other mechanisms (Bretscher, 2014; Whitelaw et al., 2017). Indeed, a recent study used a combination of traction force microscopy, quantitative RICM (reflection interference contrast microscopy), micropatterning and expansion microscopy to determine the forces and mechanisms involved in parasite gliding. Together, the data suggest a mechanism, where the MyoA motor directs the traction force, allowing transient energy storage by the subpellicular microtubule cytoskeleton and therefore sets the thrust force required for gliding (Pavlou et al., 2020). Interestingly, using expansion microscopy, it was also demonstrated that MyoA, the central motor of the glideosome is coaligned with subpellicular microtubules, arguing for a direct or indirect connection between the acto-myosin system and subpellicular microtubules. This leads to provocative questions regarding the exact location and orientation of the actomyosin system at the IMC (Tardieux & Baum, 2016).
Current models (Soldati, Foth, & Cowman, 2004; Tardieux & Baum, 2016) suggest that the acto-myosin system is localized between the IMC and the PM of the parasite. This space is very narrow (20–30 nm) and surprisingly electron lucid, indicating a low density of proteins. In contrast, just below the IMC is the so called sub-pellicular network (SPN), an electron dense network consisting of interwoven 8–10 nm filaments which gives the parasite strength and stability (Mann & Beckers, 2001). The filaments making up this network are named alveolins, a family of intermediate filament-like proteins conserved between all members of the infrakingdom Alveolata (Gould, Tham, Cowman, McFadden, & Waller, 2008). Interestingly, F-actin was demonstrated to be localized just beneath the IMC in early EM studies and more recently actin has been purified from the SPN (Patron et al., 2005; Yasuda, Yagita, Nakamura, & Endo, 1988). In good agreement, analysis of actin dynamics during motility and host cell invasion demonstrated that most of the F-actin formation occurs within the cytosol of the parasite and that filaments are subsequently transported in a retrograde fashion along the surface, probably the IMC (Del Rosario et al., 2019). Furthermore, mutants where the retrograde flow of F-actin is blocked, such as a null mutant for myoA, demonstrated a partial colocalisation of F-actin with the subpellicular microtubules (Del Rosario et al., 2019), leading to the question if the molecular organisation and localisation of the actomyosin system could face towards the cytosol of the parasite, being in direct contact with subpellicular microtubules. In support of this hypothesis, recent studies demonstrate that subpellicular microtubules are connected and stabilised by proteins that are also crucial components of the glideosome, such as GAPM-proteins (Harding et al., 2019) or GAP40 (Harding et al., 2016), indicating that they are linked to the glideosome of the parasite (Harding et al., 2019). Together, these novel findings indicate that the parasite's F-actin and microtubule system engage in extensive crosstalk to ensure parasite motility and host cell invasion (Figure 1c,d).
Furthermore, analysis of F-actin dynamics during invasion of Toxoplasma and Plasmodium (Del Rosario et al., 2019) revealed a novel role of F-actin during invasion that appears to act in parallel with the glideosome. Apicomplexan parasites establish their own entry portal, a ring-like junction through which they actively invade the host cell (Besteiro, Dubremetz, & Lebrun, 2011; Riglar et al., 2011). During this process the parasite is deformed and a recent study suggests that the host cell exerts counter-pressure on the junction, which can result in abortive invasion events, especially when components of the acto-myosin system are disrupted (Bichet et al., 2014; Bichet et al., 2016). This situation is akin to other eukaryotes, where the nucleus represents a major obstacle for the migration through a constricted environment (McGregor, Hsia, & Lammerding, 2016). When F-actin dynamics of invading parasites were analysed, a meshwork surrounding the nucleus could be detected, leading to the hypothesis that F-actin, potentially in concert with the subpellicular microtubules facilitates nuclear entry through the junction in a push-and-pull mechanism, as observed for other motile eukaryotic cells when moving through constricted environments (Del Rosario et al., 2019) (Figure 1d). Indeed, an integration of the nucleus with the cytoskeleton is observed in most eukaryotes and this is facilitated by the so-called LINC-complex. However, to date, the components of the LINC complex in apicomlexan parasites (similar to many nuclear envelope proteins) remain unknown (Rout, Obado, Schenkman, & Field, 2017).
On the other hand, the requirement o
留言 (0)