
記住我




The corticotropin-releasing factor (CRF) and the endocannabinoid (eCB) systems are important players of the negative valence system and are implicated in various stress-related psychopathologies (for reviews see Reference [1, 2]). The role of the neuropeptide CRF in the regulation of the endocrine and behavioral stress response has been well described.1 CRF secretion from neurons of the paraventricular nucleus of the hypothalamus (PVN) into the portal blood of the median eminence triggers the activation of the hypothalamic–pituitary–adrenal (HPA) axis. Within the brain, release of CRF modifies anxiety-related behavior via CRF receptor type 1 (CRFR1) in a cell type-specific manner.3, 4 Intracerebroventricular (ICV) injections and central overexpression of CRF promote anxiogenic effects in rodents that are mediated by CRFR1, likely expressed by forebrain glutamatergic neurons.4-10 Likewise, reducing CRF activity, and CRFR1 signaling via midbrain dopaminergic neurons leads to anxiolytic behavioral phenotypes.3, 4, 11, 12 Human studies have also showed a role for CRF in anxiety disorders as CRF levels are elevated in individuals suffering from post-traumatic stress disorder (PTSD).13, 14 Besides its involvement in promoting anxiety, CRF also plays a role in arousal. In rodents, increased central CRF led to increased acoustic startle responses (ASR) and decreased startle habituation.15-18
The eCB system is a retrograde messenger system modulating synaptic transmission, whereby the stimulation of the presynaptically located cannabinoid type 1 (CB1) receptor by endocannabinoids suppresses neurotransmitter release. Studies using CB1-deficient mice and pharmacological manipulations confirmed a role of the eCB system in regulating anxiety-related behaviors in rodents, again in a cell type-dependent manner. Confronted with strongly aversive stimuli, CB1 knockout mice exhibited an increased anxiety-like behavior.19 Hereby, the cellular identity of CB1-expressing neurons plays an opposing role with GABAergic neurons mediating anxiogenic and glutamatergic neurons anxiolytic effects (for review see Reference [2]). In respect to PTSD, affected individuals have been found to have reduced levels of the eCB anandamide (AEA) and upregulated CB1 receptor expression.20, 21
Interestingly, CRF and the eCB system seem to regulate fear, anxiety, behavioral and hormonal stress responses in an antagonistic manner. A considerably large body of literature describes the functional interaction of the systems with a focus on HPA axis regulation (for reviews see Reference [22, 23]). For instance, Di and colleagues described a model that suggests a glucocorticoid receptor-triggered activation of eCB synthesis on parvocellular neurons leading to a CB1-mediated inhibition of glutamate release from presynaptic neurons as a mechanism for the fast feedback inhibition of CRF by corticosterone (CORT;24 for review see Reference [25]). As another example of this interaction, CRF acts via CRFR1 to trigger fatty acid amide hydrolase (FAAH) activity within the basolateral amygdala. Activation of FAAH, the main degrading enzyme of AEA, attenuates retrograde AEA signaling thus disinhibiting glutamatergic inputs to the basolateral amygdala. The resulting activation of the pyramidal projection neurons facilitates HPA axis activation.26-33 Furthermore, the medial prefrontal cortex has been identified as a critical hub for the negative feedback inhibition of the HPA axis with a CB1-mediated cessation of the stress response.34
CRF and CB1 are co-localized in several brain regions, for example, in the piriform and prefrontal cortex, the bed nucleus of the stria terminalis (BNST), the PVN, the amygdala and the locus coeruleus.35-37 In spite of the intriguing involvement of both the CRF and eCB system in anxiety-related behaviors, nothing is known about the physiological relevance of this co-expression. Therefore, here we crossbred previously generated and validated mouse lines, the CRF-IRES-Cre38 and the CB1-floxed39 mice to generate a conditional knockout mouse line lacking CB1 specifically in CRF-positive neurons (CB1cKO-CRF). CB1cKO-CRF and wildtype littermates were assessed for anxiety and fear-related behavior, arousal, and HPA axis function. We additionally characterized the mutant mice after a traumatic experience, given the potential involvement of both CRF40, 41 and endocannabinoids in PTSD-like phenotypes (for reviews see Reference [42, 43]).
2 MATERIALS AND METHODS 2.1 AnimalsMale and female CB1cKO-CRF and CB1Ctrl (male: 2–7 months age, female: 2–6 months age) and male C57BL/6NRjMpi (originating from Janvier, 3 months age) mice were bred in the vivarium of the Max Planck Institute of Biochemistry, Martinsried, Germany. After the transfer to the animal facility at the Max Planck Institute of Psychiatry, mice were permitted a recovery period of at least 10 days before starting experiments. The animals were group-housed under standard housing conditions in Green Line IVC Sealsafe mouse cages (Tecniplast, Hohenpeißenberg, Germany) equipped with bedding and nesting material and a wooden rodent tunnel (ABEDD, Vienna, Austria). Animals had access to food and water ad libitum and were maintained in a 12/12-hours normal light/dark cycle (lights on at 6 am). Behavioral testing was performed during the light phase. All experimental procedures were approved by the Government of Upper Bavaria (Regierung von Oberbayern, 55.2-2532.Vet_02–17-206) and performed according to the European Community Council Directive 2010/63/EEC. All efforts were made to reduce the number of experimental subjects and to minimize, if not exclude, any suffering.
2.2 Generation of CB1cKO-CRF mice and genotypingCB1cKO-CRF mice originated from crossing of female CRF-IRES-Cre mice38 (Jackson Laboratory stock no: 012704) with male CB1-floxed mice39 (CB1 coding region is flanked by two loxP sites). Experimental animals were generated by breeding female heterozygous CB1cKO-CRF with male heterozygous CB1-floxed mice. Cre-positive (CB1cKO-CRF) and Cre-negative (CB1Ctrl) littermates were used for experiments. Genotyping was performed by PCR using the following primers: G50 (5′-GCTGTCTCTGGTCCTCTTAAA-3′), G51 (5′-GGTGTCACCTCTGAAAACAGA-3′), G53 (5′-CTCCTGTATGCCATAGCTCTT-3′), G100 (5′-CGGCATGGTGCAAGTTGAATA-3′), and G101 (5′-GCGATCGCTATTTTCCATGAG-3′). All animals were re-genotyped after completion of the experiment.
2.3 Single and double in situ hybridizationExpression analysis was performed on 20 μm thick coronal brain slices thaw mounted onto SuperFrost Plus slides (Thermo Fisher Scientific, Waltham, MA, USA). Single and double in situ hybridization was performed as described previously.4 The following riboprobes were used: Crf (nucleotides 70–469 of GenBank accession number NM_205769.2) and Cnr1 (nucleotides 597–2129 of NM_007726.4). For double in situ hybridization, CRF riboprobes were labeled with radioactive sulfur while CB1 riboprobes were digoxigenin (DIG)-labeled. Image analysis was performed blind of genotype using open-source Fiji image processing software.44 Gray values were measured in defined areas of the piriform cortex which are outlined in Figure 1F,G. To facilitate the reading flow, we use capitalized abbreviations for gene, mRNA and protein names throughout the manuscript.

Co-localization of CRF and CB1 in the mouse brain. (A–D) Double in situ hybridization using sulfur-labeled CRF and DIG-labeled CB1 ribo-probes on C57Bl/6 mouse brain sections (n = 2). (A) Piriform cortex, (B) Paraventricular nucleus of the hypothalamus (PVN), (C) Lateral hypothalamic area, (D) Barrington's nucleus. Gray arrowheads: CRF and CB1 double-positive cells. Brightfield images. Scale bars: 20 μm. (E) Breeding scheme. Female CRF-IRES-Cre were crossed with male CB1-floxed mice. (F, G) Single in situ hybridization using sulfur-labeled CB1 ribo-probes on male knock-out (cKO-CRF, n = 3) and wildtype (Ctrl, n = 3) mouse brain sections and measurement of gray values for the anterior (F) and posterior (G) Piriform cortex areas. Darkfield images. Scale bars: 200 μm. Same-color data points represent measurements of brain slices originating from the same mouse. **p < 0.01 (unpaired t-test)
2.4 Stress and behavioral proceduresMale and female mice were tested as different groups (i.e., on different times of the day or different days) to avoid unspecific carry-over effects by sex pheromones. We therefore do not draw conclusions on sex differences, only genotype differences.
2.4.1 Open field testThe open field test (OFT) was used to assess locomotor activity and anxiety measures. Mice were placed into a dimly lit (100 lx) square arena (W50 × L50 × H40 cm) with opaque walls facing the wall. The animals could freely explore the arena for 15 min. The movement of the mouse was video recorded using ANY-maze software (Stoelting Co., Dublin, Ireland) and the time spent in the center zone (W35 × L35 cm, 1225 cm2) versus the outer zone (W7.5 cm, 1275 cm2) and the total distance moved were analyzed. The arena was cleaned with soap and water after each trial.
2.4.2 Elevated plus mazeIn the elevated plus maze (EPM) test, mice were exposed to an elevated (32 cm above ground) plus-shaped maze consisting of two opposing arms enclosed by opaque Plexiglas walls (L27 × W5 × H14 cm) and two opposing arms (L27 × W5 cm) without walls (except for a small rim), connected by a central zone (L5 × W5 cm). After being placed in the end of one of the closed arms facing the wall, mice could freely explore the maze for 15 min. The experiment was video-recorded using ANY-maze tracking software and the time spent in the open arms and the latency to enter an open arm were determined. The setup was cleaned with soap and water after each trial.
2.4.3 Light dark boxThe light dark box (LDB) apparatus consisted of two compartments. One compartment (W20 × L29 × H25 cm) was made of white plexiglass walls and brightly lit (200 lx) while the other one was a black dark box (W20 × L15 × H25 cm). Both compartments were connected via a door (W6 × H10 cm). The animal was placed in the brightly lit compartment and was allowed to freely move between to two areas for 15 min. The experiment was videotaped by ANY-maze tracking software and the time spent in the light zone was analyzed. The number of fecal boli were counted. The arena was cleaned with soap and water after each trial.
2.4.4 Beetle Mania TaskThe Beetle Mania task (BMT) was performed essentially as described previously.45 In brief, mice were placed into the end of a gray polyethylene arena (L100 × W15 × H37 cm; 80–120 lx) and the number of rearings was scored during the first 5 min (habituation phase). Subsequently, a robo-beetle (Hexbug Nano, Innovation First Labs Inc., Greenville, TX, USA) was inserted most distantly from the mouse. For another 5 min, the number of contacts and avoidance behavior (mouse withdrew from the beetle upon contact) were scored by an experienced observer blind to genotype and experimental condition. The arena was cleaned with soap and water after each trial.
2.4.5 Acoustic startle responseIn the acoustic startle response (ASR) test, startle reflexes to acoustic stimuli were measured using the Startle Response System (TSE Systems GmbH, Bad Homburg, Germany). Animals were placed into the metal grid cage (L9.5 × W4 × H4.5 cm) sitting on the measuring platform. For the measurement of the intensity-response curve, the following protocol was applied: After a 5-min habituation period without sound presentation, white noise pulses of 70 dB(A), 90 dB(A), and 105 dB(A) (duration: 20 ms) were presented in a pseudo-randomized order 30 times each, interspersed with 18 control trials (no sound presentation). Inter-trial intervals were of 13–25 s length. The startle amplitude was defined as the peak amplitude in grams within the first 100 ms after stimulus onset. The prepulse inhibition (PPI) protocol consisted of 5 initial white noise pulses of 105 dB(A) followed by pulses of 105 dB(A) with prepulses of 75 dB(A). Prepulses were presented 50, 150 and 500 ms prior to the startle pulse. Each prepulse-pulse combination was presented 27 times in a pseudo-randomized order interspersed with 27 white noise pulses of 105 dB(A). Inter-trial intervals were of 13–25 s length. PPI was calculated as a percentage PPI = 100 × (1 – [startle response for prepulse + startle trial)/(startle response for startle stimulus alone trial]).
2.4.6 Trauma protocolFoot shock delivery and trauma memory assessment were performed as described previously.46 In brief, animals were placed into a cubic-shaped conditioning chamber (MED Associates, Fairfax, VT, USA) with a metal grid floor through which two electric foot shocks of 1.5 mA and 2 s duration were delivered. The chamber had been cleaned with 70 % ethanol. Animals of the control group underwent the same procedure without receiving a foot shock. Thirty days later, all animals were placed into a neutral context (a cylindric chamber with bedding instead of metal grid and cleaned with 1 % acetic acid) for 3 min. On the subsequent day, the mice were re-exposed to the conditioning chamber that had been cleaned with ethanol to test for conditioned fear. The trials were videotaped and freezing time and number of rearings were analyzed by an experienced observer.
2.4.7 Acute stress and plasma CORT measurementsMice were single-housed for 10 days. On testing day, the animals were restrained in a 50 ml Falcon tube (equipped with holes for tail movement and oxygen supply) for 15 min in their home cage during morning hours of the light phase. At the end of the restraint (t15), a tail cut was made at the middle part of the tail. Blood was collected in EDTA-coated tubes (Sarstedt, Nümbrecht, Germany). Blood collection was repeated 30 min (t30) and 90 min (t90) after the onset of the restraint. Trunk blood was collected 2 weeks after the restraint stress (basal) during morning hours of the light phase. All blood samples were centrifuged at 8g for 15 min at 4°C. Plasma was retrieved from the supernatant and CORT concentrations were measured using a commercially available radioimmunoassay kit (MP Biomedicals, Eschwege, Germany).
2.5 Statistical analysisData are presented as means ± SEM. For normally distributed data, unpaired t-tests, one-way analysis of variance (ANOVA) followed by Tukey's post-hoc test or two-way analysis of variance (ANOVA) for repeated measures followed by Bonferroni post-hoc analysis were performed. Kruskal-Wallis followed by Dunn's multiple comparison tests was employed for non-parametric distribution. Statistical significance was accepted if p < 0.05. All statistical analyses were performed using GraphPad Prism 9.0.
3 RESULTS 3.1 Co-localization of CRF and CB1 in the mouse brainTo assess the level of CRF and CB1 co-expressing cells in the mouse brain, we performed a double in situ hybridization on brain slices of male C57BL/6 mice. We found a moderate level of co-localization in the piriform cortex (mostly in the anterior part, Figure 1A), and the Barrington's nucleus (Figure 1D). A low level of co-localization was observed in the PVN (Figure 1B) and the lateral hypothalamic area (Figure 1C). We found no co-localization in the central and basolateral amygdala and the BNST (data not shown).
3.2 Conditional knockout of CB1 in CRF-positive neuronsTo study the effect of a potential unrestrained CRF release from CB1-positive neurons, we bred female CRF-IRES-Cre mice with male CB1-floxed mice (Figure 1E). The resulting conditional knockout mice (CB1cKO-CRF) were expected to express a reduced level of CB1 in the aforementioned brain areas compared with the wildtype littermates (CB1Ctrl). To confirm this assumption, we performed a single CB1 in situ hybridization on brain slices of male CB1cKO-CRF and CB1Ctrl mice. We measured the gray values of a defined area of the lateral anterior and posterior piriform cortex (Figure 1F, G), a region shown to harbor CFR-CB1 double-positive cells. We found a reduction of gray value in CB1cKO-CRF compared with wildtype mice in the anterior piriform cortex (t12 = 3.9, p < 0.01; Figure 1F) but not in the more posterior part of this area (t14 = 0.8, p = 0.44; Figure 1G).
3.3 Baseline behavioral characterization of CB1cKO-CRFTo assess behavioral consequences of the conditional knockout, we exposed male and female CB1cKO-CRF and CB1Ctrl mice to a battery of behavioral tests (Figure 2A). We found no significant differences between knockout and wildtype animals in a 15-min OFT, measuring the total distance traveled (males: t20 = 0.14, p = 0.89; females: t22 = 0.46, p = 0.65; Figure 2B) nor in the time spent in the inner and outer zone of the arena (males: F(1,40) = 2.25, p = 0.14; females: F(1,44) = 0.56, p = 0.46; Figure 2C). The mice were next tested in the EPM for anxiety-like behavior. No differences in the time spent in the open arms (males: U = 46, p = 0.23; females: t22 = 1.60, p = 0.12; Figure 2D) nor in the latency to enter the open arms (males: U = 41, p = 0.13; females: U = 63, p = 0.62; Figure 2E) were observed between the genotypes. Similarly, there were no differences in the time spent in the light compartment during a 15-min LDB test (males: t21 = 0.28, p = 0.78; females: t22 = 1.06, p = 0.30; Figure 2F) nor in defecation (males: t21 = 0.25, p = 0.81; females: t22 = 0.17, p = 0.86; Figure 2G).
 Baseline behavioral characterization. (A) CB1cKO-CRF and CB1Ctrl male (n = 11/12) and female (n = 12/12) mice were exposed to a behavioral test battery for baseline characterization. d, day. Distance moved (B) and time in the inner (IZ) and outer zone (OZ, C) in the open field test (OFT). Time spent in the open arms (D) and latency to the first open arm entry (E) in the elevated plus maze test (EPM). Time spent in the light zone (F) and defecation (G) during the light–dark box test (LDB). Number of rearing events (H) and avoidance behavior as a fraction of total contacts with the robo-beetle (I) during the Beetle Mania Task (BMT).45
Baseline behavioral characterization. (A) CB1cKO-CRF and CB1Ctrl male (n = 11/12) and female (n = 12/12) mice were exposed to a behavioral test battery for baseline characterization. d, day. Distance moved (B) and time in the inner (IZ) and outer zone (OZ, C) in the open field test (OFT). Time spent in the open arms (D) and latency to the first open arm entry (E) in the elevated plus maze test (EPM). Time spent in the light zone (F) and defecation (G) during the light–dark box test (LDB). Number of rearing events (H) and avoidance behavior as a fraction of total contacts with the robo-beetle (I) during the Beetle Mania Task (BMT).45
To assess active versus passive fear responses to a potentially threatening stimulus, the animals were subjected to the BMT. We could not show any differences in the number of rearing events during the baseline period (males: t21 = 0.12, p = 0.90; females: t22 = 0.00, p > 0.99; Figure 2H). Also, upon confrontation with the robo-beetle, there was no significant difference in the avoidance behavior between the two genotypes (males: t21 = 1.57, p = 0.13; females: t22 = 1.28, p = 0.21; Figure 2I).
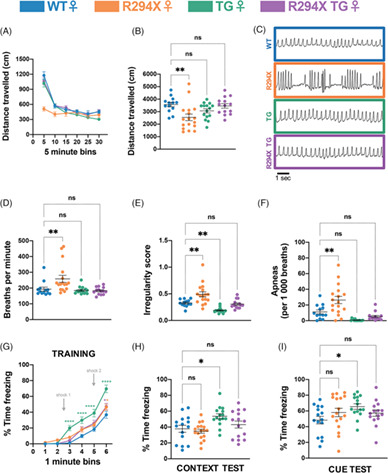
A new cohort of male and female CB1cKO-CRF and CB1Ctrl mice was tested for the ASR and for circulating CORT levels (Figure 3A). We exposed mice to white noise pulses of different intensity and measured the startle amplitude. As showed by 2-way ANOVA, there was a significant interaction between genotype and startle pulse intensity in male mice (F3,69 = 4.47, p < 0.01; Figure 3B) with post-hoc analysis confirming a higher startle amplitude of CB1cKO-CRF at the highest white noise intensity (105 dB(A)) compared with wildtype controls. This effect could not be observed in female mice (F3,66 = 0.16, p = 0.92; Figure 3B). Next, mice were tested in a PPI task. No significant difference in the startle response could be shown between CB1cKO-CRF and wildtype mice (males: F2,46 = 2.07, p = 0.14; females: F2,44 = 0.59, p = 0.56; Figure 3C). After sufficient time for recovery, we measured basal and post-stress blood plasma CORT concentrations. Mice were restrained for 15 min and blood was sampled via a tail cut at the end of the restraint period (t15), as well as 30 (t30) and 90 (t90) min after the beginning of the stressor. Mice showed an increase in plasma CORT after the restraint, but there was no significant difference between the genotypes (males: F3,69 = 0.75, p = 0.53; females: F3,63 = 0.48, p = 0.70; Figure 3D).

Startle response and CORT measurements. A new cohort of male and female CB1cKO-CRF and CB1Ctrl mice was tested for acoustic startle responses (ASR; nmale = 13/12, nfemale = 12/12) and plasma corticosterone (CORT; nmale = 11/15, nfemale = 11/12) levels. d: day (A). Intensity-response relationship between sound-pressure levels (SPL) of the white noise pulses and ASR (B). Prepulse inhibition (PPI) with different inter-pulse intervals (IPI, C). CORT blood plasma concentrations at basal and 15, 30, and 90 min after restraint stress (D). **p < 0.01 (two-way rm-ANOVA followed by Bonferroni's post-hoc test)
3.4 Behavioral characterization of CB1cKO-CRF after trauma incubationBoth the CRF and CB1 systems are known to be involved in trauma-related behavioral changes. Therefore, we exposed experimentally naïve male and female CB1cKO-CRF and wildtype controls to a trauma protocol. Mice were assigned to four experimental groups per sex, following a 2 × 2 design (genotype x trauma). All groups were placed into a shock chamber, with two groups receiving two electric foot shocks of 1.5 mA (cKO-S+ and Ctrl-S+), while the other two groups remained non-shocked (cKO-S- and Ctrl-S-). Four weeks later, we assessed generalized trauma-associated fear followed by measurement of active and passive fear responses (Figure 4A). Mice that had received a foot shock froze significantly more (Kruskal-Wallis test; males: p < 0.0001; females: p < 0.0001; Figure 4B) and exhibited significantly fewer rearing events than non-shocked controls upon exposure to a neutral test context (Kruskal-Wallis test; males: p < 0.0001; females: p < 0.0001; Figure 4C). These observations were independent of genotype or sex. The same findings for freezing and rearing were shown when the mice were re-exposed to the shock context on the next day (Kruskal-Wallis test; males: p < 0.0001; females: p < 0.0001; Figure 4D,E).

Behavioral characterization after trauma exposure. (A) Male (n = 8–14 per group) and female (n = 8–14 per group) CB1cKO-CRF and CB1Ctrl mice received two electric foot shocks of 1.5 mA (cKO-S+ and Ctrl-S+) or were assigned to the non-shocked control group (cKO-S- and Ctrl-S-). After an incubation time of 30 days, generalized and contextual fear were assessed in a novel (cylinder) and in the shock context (chamber), respectively. One week later, mice were exposed to the Beetle Mania Task (BMT). Time spent freezing (B) and number of rearing events (C) during exposure to the novel environment. Time spent freezing (D) and number of rearing events (E) during re-exposure to the shock environment. Number of rearing events (F) and avoidance behavior as a fraction of total contacts with the robo-beetle (G) during the BMT. **p < 0.01, ***p < 0.001, ****p < 0.0001 (1-way ANOVA for B-E, G; Kruskal-Wallis test for F)
Next, we tested the mice in the BMT to measure the consequences of trauma on active versus passive fear responses. During the baseline period without robo-beetle, cKO-S+ mice showed significantly reduced vertical exploration behavior (rearing events) compared with cKO-S- (Kruskal-Wallis test; males: p < 0.01; females: p < 0.001) but there was no significant difference between CB1cKO-CRF and CB1Ctrl (Figure 4F). Avoidance behavior in response to the robo-beetle was unchanged between genotypes and trauma experience (males: F3,39 = 0.88, p = 0.46; females: F3,40 = 2.09, p = 0.12; Figure 4G).
4 DISCUSSIONThe current study investigated consequences of the selective deletion of CB1 receptors from CRF neurons by means of a newly generated conditional knockout mouse line (CB1cKO-CRF). The cell-type specific lack of CB1 expression caused an increase in the ASR in male but not female mice with no consequences on locomotion, anxiety- and fear-related behavior and HPA axis activity. Likewise, no behavioral differences were found following trauma exposure.
In wildtype mice, we could show the co-localization of CB1 and CRF mRNA in the piriform cortex, the PVN, the lateral hypothalamic area, and the Barrington's nucleus. This is in agreement with previous studies, which additionally report co-expression in the prefrontal cortex, the BNST, and the amygdala.35, 36 An immunohistochemical approach showed CB1 and CRF protein co-localization in the locus coeruleus in axon terminals of neurons projecting from the central amygdala (CeA).37 Our failure to show co-localization in the amygdala might be ascribed to differences in the sensitivity of the methods and/or generally low levels of expression of both CB1 and CRF under basal conditions.
To study the consequences of CB1 loss on CRF-expressing neurons, we crossed CRF-IRES-Cre38 and CB1-floxed39 mouse lines. Both mutant mouse lines have been well investigated and successfully used for knockout and mapping studies.3, 47-50 Using in situ hybridization, we could confirm a reduction of CB1 for the piriform cortex of CB1cKO-CRF. Even though we did not perform co-expression analyses in mutant mice due to the low abundant expression of both CRF and CB1, it is highly likely that this reduction results from the selective deletion of CB1 expression in CRF-positive neurons.
In order to study behavioral consequences of the knockout on the negative valence system, we exposed male and female mutants and their respective wildtype littermate controls to a variety of behavioral paradigms. We found no difference in locomotor activity or fear and anxiety-related behaviors between CB1cKO-CRF and controls of both sexes. If we assume that the lack of CB1 receptors may result in unrestrained release of CRF, we would have expected increased locomotion and anxiety.6, 9, 10, 51-54
Male CB1cKO-CRF showed an increased startle amplitude compared with male littermate controls. This is in line with previous studies reporting increased ASR and decreased startle habituation following ICV administration of CRF or overexpression of CRF.15-17, 55, 56 The lack of an arousal effect in female CB1cKO-CRF could be attributed to sex dimorphisms of the CRF system, as has been shown in terms of behavior, HPA axis function, and gene expression17 (for reviews see Reference [57, 58]). We can only speculate about the pathway involved in the hyperarousal shown by male mutants. There is the likelihood of an involvement of projections from the CeA to the BNST.18 A contribution of CRF signaling within the CeA for the generation of active versus passive fear59 however, appears to be less likely, given the lack of effects in the BMT, a test designed to measure unconditioned active versus passive fear.45, 60, 61 Besides, we could not reproduce the previously described co-localization of CRF and CB1 within the CeA.
Under basal conditions, our experiments showed no effects from the knockout on fear and anxiety. This might be due to a lack of activation of the CRF system under the experimental conditions. Exposure to a stressor has been found to increase CRF expression in various brain regions,62 and it has been suggested that endogenous CRF release must be triggered by a stressor prior to a paradigm in order to observe behavioral effects.63 To challenge the CRF system, we exposed the mice to both an acute restraint followed by measurement of HPA axis activity and assessed behavior after incubation of a traumatic event.46 Despite the well-described regulation of CB1 on the HPA axis, we failed to detect differences in plasma CORT levels between the genotypes, both at basal and post-stress time points. Likewise, no behavioral changes were observed after trauma exposure.
The very distinct behavioral phenotype of male CB1cKO-CRF might be explained in several ways. We observed an overall low level of co-localization of CRF and CB1 throughout the brain, that is, only a low number of cells is affected by the knockout. Next, it is unclear whether these cells are activated at all during the tasks employed and whether enough CRF is released under basal conditions. Another important question is whether CRF release is even under control of CB1. Depolarization-induced suppression of excitation and inhibition are well-established concepts of CB1-controlled release of the neurotransmitters glutamate and GABA, respectively.64, 65 Less is known about CB1 inhibiting the release of neuropeptides. A functional interaction of CB1 and the cholecystokinin (CCK) system in regulating fear memory has been shown.66, 67 But while CCK and CB1 are very highly co-expressed for example in hippocampal basket cells, we observed an overall low abundant co-expression of CRF and CB1. Moreover, the possibility of neurotransmitter co-transmission must be borne in mind. CRF is mostly expressed in GABAergic neurons in the brain,68, 69 with the exception of the piriform cortex and the PVN where it is expressed in glutamatergic neurons.70, 71 Thus, whether the observed hyperarousal of male CB1cKO-CRF can be attributed to disinhibited release of CRF or neurotransmitters remains to be confirmed using CRFR1 or CRFR2 antagonists or new genetic models.
In conclusion, we show a selective increase in the ASR of male mice lacking CB1 receptor expression in CRF neurons with no alterations in the negative valence system and HPA axis activity.
ACKNOWLEDGMENTSWe thank Andrea Ressle and the Scientific Core Unit for Genetically Engineered Mouse Models for performing the genotyping of the mouse line. We would like to thank Dr. Jessica Keverne for proofreading of the manuscript.
Julia Ruat was supported by the International Max Planck Research School for Translational Psychiatry (IMPRS-TP). Alice Hartmann received a fellowship from Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP; BEPE-2018/17387-9). Daniel E. Heinz was supported by the Federal Ministry of Education and Research (BMBF) and the Max Planck Society. Alon Chen is the incumbent of the Vera and John Schwartz Professorial Chair in Neurobiology at the Weizmann Institute of Science; the Head of the Max Planck Society–Weizmann Institute of Science Laboratory for Experimental Neuropsychiatry and Behavioral Neurogenetics gratefully funded by the Max Planck Foundation; and the Head of Ruhman Family Laboratory for Research in the Neurobiology of Stress at the Weizmann Institute of Science. This work is supported by Bruno and Simone Licht (AC) and Roberto and Renata Ruhman. This study was further supported by the German-Israeli Foundation for Scientific Research and Development (GIF; I-1442-421. 13/2017) to Carsten T. Wotjak.
CONFLICT OF INTERESTThe authors declare no conflict of interest.
留言 (0)