
記住我
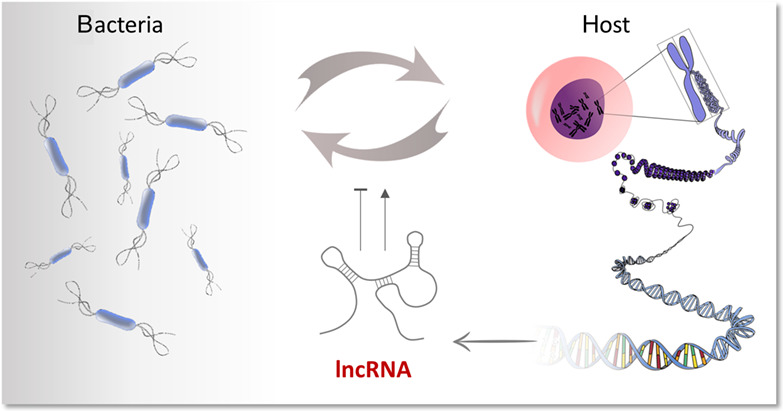
The importance of gene regulation by RNA for bacterial competitiveness and survival is well established, but new facets and even new mechanisms are still emerging. In transcriptomic studies, usually steady-state RNA levels are determined, which result from the rates of transcription and decay. Bacterial mRNAs are highly unstable, with typical half-lives within the range of few minutes. Furthermore, the mRNA half-lives change with changing environmental conditions (Bernstein et al., 2002; Klug, 1991; Nilsson et al., 1984; Vargas-Blanco et al., 2019). Such stability changes are often directly or indirectly caused by stimuli-induced regulatory RNAs (Adnan et al., 2015; Durand et al., 2015; Lenz et al., 2004; Pfeiffer et al., 2007) or by sensory RNA elements (Nou & Kadner, 1998; Richards & Belasco, 2021b). In addition, regulatory RNAs influence the initiation or the progress of mRNA transcription (Bossi et al., 2012; Sedlyarova et al., 2016; Silva et al., 2019; Wassarman & Storz, 2000).
Regulation with RNA (riboregulation) is widespread, mechanistically diverse, and affects virtually all aspects of the bacterial life. Among others, it helps bacteria to adapt to stress conditions (Adnan et al., 2015; Altuvia et al., 1997; Augagneur et al., 2020; Lalaouna, Baude, et al., 2019; Morita et al., 1999; Nocker et al., 2001), maintain metabolic homeostasis (Hoyos et al., 2020; Khan & Görke, 2020; Papenfort et al., 2013; Wilderman et al., 2004), regulate photosynthesis genes (Dühring et al., 2006; Georg et al., 2014; Mank et al., 2012), remodel gene expression during growth (Majdalani et al., 1998; Sedlyarova et al., 2016; Wassarman & Storz, 2000), produce biofilms (Holmqvist et al., 2010; Liu et al., 2016; Papenfort et al., 2015; Schoenfelder et al., 2019), coordinate expression of virulence factors (Boisset et al., 2007; Brewer, Twittenhoff, et al., 2021; Eisenbart et al., 2020; Jia et al., 2021; Pappesch et al., 2017; Twittenhoff, Heroven, et al., 2020; Westermann et al., 2016), and to survive antibiotic treatment (Borgmann et al., 2018; Peschek et al., 2020; Zhang et al., 2017). Numerous excellent and comprehensive reviews on prevalence, mechanisms, and evolution of regulatory RNAs are available (e.g., Adams & Storz, 2020; Babitzke et al., 2019; Carrier et al., 2018; Chen, Morita, & Gottesman, 2019; Dersch et al., 2017; Felden & Cattoir, 2018; Hör, Matera, et al., 2020; Ignatov & Johansson, 2017; Santiago-Frangos & Woodson, 2018; Turnbough, 2019; Wagner & Romby, 2015; Waters & Storz, 2009). Here, after a general overview of the principles of RNA regulation in bacteria, I will focus on new, unexpected functions of the conserved trp attenuator in Gram-negative bacteria, which indicate emerging mechanisms in response to antibiotics exposure.
2 PROTEIN-BINDING, TRANS-ACTING SRNASRegulatory RNA, which acts independently of its genomic locus, is trans-acting RNA. In bacteria, many small RNAs (sRNAs), typically ranging from 50 to 300 nucleotides, act in trans (Mizuno et al., 1984; Zhang et al., 2003). They have two principle modes of action: protein binding or base pairing.
The prime example for a protein-binding sRNA is 6S RNA. In Escherichia coli, it accumulates throughout growth, binds the sigma70-RNA polymerase (RNAP) holoenzyme, and contributes to the changes in gene expression in the stationary phase (Cavanagh et al., 2008; Wassarman & Storz, 2000). 6S RNA is highly conserved and some bacteria have two or more 6S RNA genes (Wehner et al., 2014). The two Bacillus subtilis 6S RNA paralogs 6S-1 and 6S-2 have different expression profiles during growth and 6S-1 seems to be functionally similar to E. coli 6S RNA. Using an undomesticated B. subtilis strain, it was shown that they both are important for the bacterial fitness under harsh environmental conditions (Thüring et al., 2021). In Bacillus, 6S-1 RNA contributes to sporulation (Cavanagh & Wassarman, 2013; Li, Zhu, et al., 2020). In anoxygenic photosynthetic bacteria, 6S RNA is important for salt stress tolerance (Elkina et al., 2017) and in cyanobacteria, for recovery from nitrogen depletion (Heilmann et al., 2017). Interestingly, in Mycobacterium smegmatis, the sRNA Ms1, which is not related to 6S RNA, interacts with the core RNAP and influences its amount (Šiková et al., 2019). Since also natural RNA aptamers were found to regulate transcription by interacting with RNAP in E. coli (Sedlyarova et al., 2017), it is possible that RNAP-binding riboregulators still remain to be discovered in other bacteria.
Other functions of protein-binding sRNAs are to sequester transcription factors or posttranscriptional regulators. Such protein-sponging functions are important for bacterial virulence. For example, B12-riboswitch-regulated sRNAs sequester the two-component response regulator EutV in Listeria and Enterococcus (DebRoy et al., 2014; Mellin et al., 2014). In E. coli, Erwinia, and Yersinia, the CsrB/RsmB sRNA and its paralogs sequester the global regulator and RNA-binding protein (RBP) CsrA/RsmA, which otherwise represses translation of genes for glycogen biosynthesis, extracellular enzymes, or transcription factors (Heroven et al., 2012; Liu et al., 1997). Furthermore, in E. coli, the sRNA GlmY titrates the RNA-processing factor RapZ away from its regular target, the sRNA GlmZ. The GlmZ sRNA is responsible for posttranscriptional activation of glmS, which is needed for cell envelope synthesis (Göpel et al., 2013; Khan et al., 2020). Protein-binding sRNAs have been much less investigated than base-pairing sRNAs, probably because not much is known about RBPs in bacteria. New high-throughput approaches such as Grad-seq (Hör, Di Giorgio, et al., 2020; Smirnov et al., 2016) and organic phase separation (Smith et al., 2020; Urdaneta et al., 2019) will certainly unravel novel RBPs and protein-binding riboregulators.
3 BASE-PAIRING, TRANS-ACTING SRNAS 3.1 Basic principlesBase-pairing, trans-acting sRNAs affect translation and/or stability of mRNA in a positive or, more often, in a negative way. Inhibition of translation is often due to binding of an sRNA to the ribosome binding site (RBS) as described for the OxyS sRNA and fhlA mRNA (Altuvia et al., 1998), but translation inhibition was also observed upon sRNA binding more upstream in the 5′-UTR (Andreassen et al., 2018; Sharma et al., 2007), or even in the coding sequence (Bouvier et al., 2008). In addition to blocking the ribosome access by the sRNA (Maki et al., 2008) several other translation inhibition mechanisms were deciphered in E. coli: The RyhB sRNA downregulates fur expression by blocking a short, upstream open reading frame (uORF), whose translation is coupled to that of the fur gene (Vecerek et al., 2007). The OmrA and OmrB sRNAs inhibit the formation of translation-activating stem-loops in the coding regions of fepA and bamA mRNAs (Jagodnik et al., 2017). Translation of manX mRNA is repressed by the RNA chaperone Hfq, and this interaction is supported by the SgrS and DicF sRNAs (Azam & Vanderpool, 2018). Finally, the SgrS sRNA interferes with the interaction between a translation enhancer and ribosomal protein S1 of manY mRNA (Azam & Vanderpool, 2020). Translation inhibition by an sRNA is often accompanied by degradation of the ribosome-free RNA (Deana & Belasco, 2005).
In bacteria, trans-acting sRNAs mostly bind their mRNA targets via imperfect complementarity, with only limited sRNA–mRNA contacts (Argaman & Altuvia, 2000; Poddar et al., 2021). Probably therefore, efficient sRNA–mRNA binding is often mediated by a proteinaceous RNA-chaperone such as Hfq (homolog of the eukaryotic Sm proteins) or ProQ (Attaiech et al., 2016; Chaulk et al., 2011; Møller et al., 2002; Smirnov et al., 2016). The mechanism of the Hfq hexamer was intensely studied, mainly in E. coli. It binds RNA via three distinct areas and according to the binding pattern, sRNAs can be divided into two classes. The majority of the studied sRNAs belong to Class I. They bind with their Rho-independent terminator to the proximal face of Hfq and with 5′ or internal regions to its arginine-rich rim, while their mRNA targets bind to the distal surface of the chaperone. Upon RNA rearrangement, annealing of the sRNA and the mRNA takes place at the Hfq rim. In contrast, Class II sRNAs bind to the proximal pore and the distal surface, and their targets bind to the rim of Hfq (Schu et al., 2015). Unstructured Hfq regions at the N- and C-terminus contribute to its RNA chaperone activity (Santiago-Frangos & Woodson, 2018).
Hfq interacts with the degradosome-organizing endoribonuclease RNase E via sRNA (Bruce et al., 2018). In this way, Hfq-dependent sRNAs recruit the RNA-degrading machinery and destroy their targets (Lalaouna, Morissette, et al., 2015; Morita et al., 2005). Thereby, the sRNA and the mRNA are co-degraded (Massé et al., 2003), or only the mRNA is degraded and thus the sRNA acts catalytically (Feng et al., 2015; Overgaard et al., 2009). Since the single-strand-specific RNase E has a monophosphate-binding pocket, it was proposed that a processed sRNA having a 5′ monophosphate more efficiently recruits RNase E (Bandyra et al., 2012), and that this works even without the help of an RNA chaperone (Baek et al., 2019). However, in vivo evidence for this mechanism remains to be provided.
The binding of an sRNA to an mRNA often results in an endonucleolytic cleavage of the mRNA. Besides RNase E, the double-strand specific RNase III can cleave the sRNA–mRNA duplex in a mechanism resembling RNAi in eukaryotic cells. The latter is exemplified by the downregulation of the porin-encoding ompA by the sRNA MicA (Viegas et al., 2011). However, since the double-strand binding domain of RNase III optimally binds to duplexes in the length of approximately 20 bp (Nicholson, 2014), RNase III-dependent cleavage was rarely described in conjunction with trans-acting sRNAs and seems to be more often used in regulation by cis-transcribed antisense RNAs (asRNAs) (Oliva et al., 2017; Vogel et al., 2004; Wen et al., 2014).
The fewer examples of positive mRNA regulation include sRNA base paring which alters an mRNA secondary structure to expose previously hidden RBS (Majdalani et al., 1998; Urban & Vogel, 2008). Additionally, sRNA binding can mask an RNase E cleavage site and increase the mRNA stability (Chen, Previero, & Deutscher, 2019; Fröhlich et al., 2013). Moreover, an sRNA can stabilize an intermediate of RNase E-dependent mRNA decay and thus selectively activate gene expression (Papenfort et al., 2013; Papenfort & Vanderpool, 2015).
Trans-acting sRNAs can base-pair not only with mRNAs, but also with other regulatory sRNAs. In such cases, one of the sRNAs acts as a “sponge” and sequesters its sRNA target and/or downregulates its level (Acuña et al., 2016; Lalaouna, Carrier, et al., 2015). Recently, in several studies, identification of sRNAs and their mRNA or sRNA base pairing partners was boosted by using high-throughput methods: Affinity purification of MS2-tagged sRNA together with interacting molecules followed by RNA sequencing (MAPS) (Carrier et al., 2016); in vivo UV crosslinking, coimmunoprecipitation, and sequencing of RNA interacting with 3 × FLAG-ProQ (CLIP-seq; Holmqvist et al., 2018); RIL-seq (RNA interaction by ligation and sequencing) of interacting RNA–RNA pairs associated with Hfq and ProQ (Melamed et al., 2016, 2020); UV cross-linking, ligation and sequencing of hybrids (CLASH) co-purified with RNase E or Hfq (Iosub et al., 2020; Waters et al., 2017); and RNA-Seq of sRNAs that are derived from 5′-UTRs or are internal to ORFs (Adams et al., 2021).
Regarding the genomic sRNA loci, the first systematic searches were focused on non-annotated regions between ORFs, and therefore in early studies, mostly sRNAs having own promoter and transcription terminator were detected (Argaman et al., 2001). Later, many 3′-UTR-derived sRNAs were identified (Chao et al., 2012), while sRNAs originating from 5′-UTRs or internal to ORFs were rarely described (Guo et al., 2014; Loh et al., 2009; Thomason et al., 2019). This currently changes with the application of Term-seq detecting 3′ ends of transcripts and of multilayered RNA-Seq approaches (Adams et al., 2021; Dar et al., 2016; Dar & Sorek, 2018).
Most of the base-pairing sRNAs are non-coding, but several dual-function sRNAs harboring small, functional ORFs were described (Gimpel & Brantl, 2017). A well-known example is the E. coli SgrS sRNA, which encodes the small protein SgrT. The base pairing of SgrS to at least nine mRNAs and the inhibition of the glucose transporter PtsG by SgrT are mechanistically independent, but both counteract the glucose-phosphate stress, which induces SgrS production (Bobrovskyy et al., 2019; Lloyd et al., 2017; Vanderpool & Gottesman, 2004). In Staphylococcus aureus, the multifaceted sRNA RNAIII, which has a length 514 nt and is one of the largest trans-acting riboregulators in bacteria, represses or activates several mRNA targets by base-pairing, and encodes the hemolytic toxin Hld (Bronesky et al., 2016; Novick et al., 1993).
3.2 Regulatory networks of sRNAsThe imperfect complementarity between sRNAs and their targets also accounts for the capability of certain sRNAs to regulate multiple genes, as shown for the Spot42, GcvB, and RybB sRNAs in E. coli (Lalaouna, Eyraud, et al., 2019; Storz et al., 2011). To base-pair with different mRNAs, an sRNA can use the same seed region as described for the 5′ target recognition domain of RybB in Salmonella (Papenfort et al., 2010). However, sRNAs also use separate modules for binding to different mRNAs, as shown for RNAIII in S. aureus (Bronesky et al., 2016), AbcR1 sRNA in Agrobacterium tumefaciens (Overlöper et al., 2014), and GcvB in E. coli (Lalaouna, Eyraud, et al., 2019). Furthermore, an sRNA can affect its multiple targets by different mechanisms. For example, the Vibrio harveyi Qrr3 sRNA deploys four distinct mechanisms to regulate quorum sensing: catalytic degradation, coupled degradation, repression through sequestration, and activation by revealing a ribosome binding site (Feng et al., 2015).
On the other hand, specific, key mRNAs can be targeted by several differently regulated sRNAs. For example, the enterobacterial curli activator mRNA csgD, which is important for biofilm formation, is regulated by no less than seven sRNAs, which relay signals such as carbon source availability, osmotic or envelope stress, and amino acid metabolism (Andreassen et al., 2018; Boehm & Vogel, 2012). Another prominent example is rpoS, which encodes the general stress sigma factor in Gammaproteobacteria. In E. coli, it is upregulated by the low-temperature activated sRNA DsrA, the cell-envelope stress-related sRNA RprA, and a processed form of the energy limitation-activated sRNA ArcZ. These sRNAs act by base-pairing within the 5′-UTR of the rpoS mRNA (McCullen et al., 2010; Mika & Hengge, 2014). Recently, it was reported that in E. coli, the CyaR sRNA downregulates rpoS expression by base pairing next to the ArcZ binding site. Additionally, the short, active form, or ArcZ binds CyaR and promotes its degradation by RNase E. Thus, ArcZ upregulates rpoS expression by two different mechanisms, mRNA activation, and sRNA sponging (Kim & Lee, 2020). Furthermore, the rpoS expression in E. coli is downregulated by the oxidative-stress induced sRNA OxyS by a still unknown mechanism (Zhang et al., 1998). Moreover, in enterohemorrhagic E. coli, rpoS is regulated by StsX, a processed sRNA, which is derived from a constitutively initiated and constitutively terminated Shiga toxin transcript (Sy et al., 2020). In Legionella, Pseudomonas, and Vibrio, rpoS is also regulated by several base-pairing sRNAs (Han & Lory, 2021; Lu et al., 2018; Saoud et al., 2021; Thi Bach Nguyen et al., 2018).
The capability of a single sRNA to bind multiple mRNAs and vice versa, and the sRNA regulation of genes encoding sigma factors and other transcription regulators result in complex sRNA networks (Bossi & Figueroa-Bossi, 2016; Nitzan et al., 2017; Storz et al., 2011). In these networks, an mRNA target can compete with other mRNAs for a shared sRNA regulator (Miyakoshi et al., 2015) and a sponge-sRNA with mRNAs for binding to sRNAs or RBPs (Lalaouna, Carrier, et al., 2015; Liu et al., 1997; Sonnleitner & Bläsi, 2014). Furthermore, many sRNAs compete for binding to the global RNA chaperones Hfq and ProQ. A recent study analyzing RNA–RNA pairs associated with Hfq and/or ProQ in E. coli revealed that they generally bind to different sRNAs, but share a set of overlapping interactors, for example, a RybB–RbsZ sRNA–sRNA pair. It was suggested that when this pair is bound to Hfq, the RbsZ sponge downregulates RybB, while binding to ProQ protects RybB from degradation (Melamed et al., 2020).
In enterobacteria, many sRNAs are functionally dependent on Hfq, the disordered C-terminal domain of which regulates the RNA competition and ensures the release of double-stranded RNA complexes (Santiago-Frangos et al., 2016). The abundance and the Hfq-binding kinetics of the particular sRNAs determine the hierarchy in their Hfq occupancy, which is important for the regulatory outcome (Faigenbaum-Romm et al., 2020; Kwiatkowska et al., 2018). By acting as a decoy for Hfq, certain sRNAs could prevent the action of other sRNAs by limiting their access to Hfq (Moon & Gottesman, 2011) or by preventing an Hfq-dependent mRNA remodeling needed for base pairing with the sRNA (Hoekzema et al., 2019). Alternatively, an sRNA can displace Hfq from its mRNA target (Park et al., 2021). Furthermore, sRNA binding to Hfq probably affects its interactions with asRNA (Bilusic et al., 2014), rRNA (Andrade et al., 2018), tRNA (Lee & Feig, 2008), and DNA (Malabirade et al., 2017), and thus its multiple functions in the cell (Dos Santos et al., 2019). Altogether, dynamic interactions, competition, displacement, and sponging determine the cross-talk between subregions of the sRNA networks and finally the posttranscriptional regulation of a specific gene in the cell.
3.3 Small RNAs in diverse bacteriaThe majority of the above sRNA examples originate from Gammaproteobacteria. In the last years, it became clear that similar mechanisms also operate in other bacteria, but certain differences were also observed. This is a fast-growing research field (Adams & Storz, 2020) and only few examples can be mentioned here.
Most Alphaproteobacteria have Hfq but not ProQ (Olejniczak & Storz, 2017). In vitro studies and heterologous expression in E. coli suggested that Hfq of the cell-cycle model organism Caulobacter crescentus is functionally similar to its E. coli homolog. However, the involvement of C. crescentus Hfq in regulation by base pairing sRNAs was not shown experimentally (Fröhlich & Velasco Gomariz, 2021; Santiago-Frangos et al., 2019). In the soil-dwelling plant symbiont Sinorhizobium meliloti, the sRNAs AbcR1 and AbcR2 are Hfq-dependent (Torres-Quesada et al., 2013). They are conserved in related pathogens belonging to Agrobacterium and Brucella, where they target multiple mRNAs (Overlöper et al., 2014; Sheehan & Caswell, 2017; Wilms et al., 2011). However, other S. meliloti sRNAs such as EcpR1 and rnTrpL (alias RcsR1), which bind to the RBS of their mRNA targets and destabilize them in an RNase E-dependent manner, do not need Hfq (Robledo et al., 2015; Robledo, García-Tomsig, & Jiménez-Zurdo, 2020). In S. meliloti, Hfq binds only a minor sRNA fraction and interacts predominantly with mRNAs (Torres-Quesada et al., 2014). To find novel RBPs in S. meliloti, recently pull-down with MS2-tagged sRNAs was conducted. Surprisingly, the S-adenosylmethionine synthetase MetK was identified as an RBP associated with functionally diverse sRNAs (Robledo, García-Tomsig, Matia-González, et al., 2021). Furthermore, in the facultative phototrophic species Rhodobacter sphaeroides, recently, the DUF1127 protein CcaF1 was described as a globally acting RBP, which influences the stability of sRNAs (Grützner, Billenkamp, et al., 2021). Currently, R. sphaeroides is the alphaproteobacterial species with the highest number of functionally characterized sRNAs. It harbors orphan, 3′-UTR and 5′-UTR derived, Hfq-dependent and Hfq-independent sRNAs, which mostly act in conjunction with RNase E (Adnan et al., 2015; Billenkamp et al., 2015; Eisenhardt et al., 2018; Grützner, Remes, et al., 2021; Hess et al., 2014; Mank et al., 2012; Müller et al., 2016; Peng et al., 2016).
In the pathogenic Helicobacter pylori (Epsilonproteobacteria), which lacks Hfq and ProQ, highly variable simple sequence repeats (SSRs) contribute to the regulation of virulence genes by phase-variation. A SSR in the 5′-UTR of the target mRNA encoding the chemotaxis receptor TlpB was found to regulate the mode of action of the RepG sRNA. This sRNA harbors a C/U rich terminator loop region, which base pairs with a homopolymeric G-repeat in the tlpB leader. Depending on the length of the G-repeat in different H. pylori strains, tlpB translation is affected differently by RepG (Pernitzsch et al., 2014). Furthermore, a variable thymine stretch in the promoter of the H. pylori sRNA NikS determines its phase-variable transcription and thus the posttranscriptional regulation of major virulence factors (Eisenbart et al., 2020).
In cyanobacteria, the sRNA AcpZ, which is derived from the 3′-end of a phycobilisome operon, downregulates the expression of the orange carotenoid protein by binding to the first codons of its ORF, thus mediating a reverse regulation of light harvesting and photoprotection (Zhan et al., 2021). Also in cyanobacteria, the iron-responsive sRNA IsaR1 has at least 15 direct target mRNAs and deploys a RBS-binding mechanism (Georg et al., 2017). However, the role of the cyanobacterial Hfq homolog is related rather to type IV pilus assembly than to sRNA metabolism (Bøggild et al., 2009; Schuergers et al., 2014; Yegorov et al., 2021), and other RNA chaperones were not described yet (Riediger et al., 2021).
The power of RNA-Seq enabled the transcriptome-wide detection and the subsequent functional analysis of sRNAs not only in well-established Gram-negative model bacteria, but also in emerging species with potentially important roles. For example, this approach led to the discovery of the sRNA donS, which influences 15 polysaccharide utilization loci in Bacteroides, a major component of the human gut microbiota. Thus, this sRNA controls 30% of the polysaccharide utilization loci, which are important for the predominance of this genus (Cao et al., 2016). Furthermore, very recently, the RNA landscape of five cancer-associated Fusobacterium strains, which lack CsrA, Hfq, and ProQ, was analyzed at different growth stages, and an oxygen-induced sRNA regulating a major porin gene was discovered (Ponath et al., 2021).
In the Gram-positive pathogen Streptococcus pyogenes, many sRNAs were detected by transcriptomics and more recently by Grad-seq (Hör, Garriss, et al., 2020; Le Rhun et al., 2016; Patenge et al., 2012). Several sRNAs were found to regulate competence and virulence genes, among them FasX, which targets multiple mRNAs (Danger et al., 2015; Hör, Garriss, et al., 2020; Pappesch et al., 2017). The virulence of the related pathogen Streptococcus pneumoniae is also controlled by sRNAs (Mann et al., 2012). In Staphylococcus, which also belongs to Bacilli, the large sRNA RNAIII uses several regulatory domains to target multiple mRNAs encoding transcriptional regulators, major virulence factors, and metabolic enzymes. Upon base-pairing with an mRNA, RNAIII mediates translation inhibition and/or RNase III cleavage, or activates gene expression by RBS liberation and/or mRNA stabilization (Bronesky et al., 2016). In S. aureus, the RsaA sRNA blocks translation of the global transcriptional regulator MgrA by imperfect base pairing with the Shine–Dalgarno sequence and by an additional loop–loop interaction within the coding region of its mRNA. It was proposed that this sRNA supports commensalism (Romilly et al., 2014). Furthermore, in the same species, the glucose limitation-activated sRNA RsaI uses two distinct base-pairing domains: a CU-rich unpaired region of RsaI binds to the RBS or the 3′-UTR of mRNAs, thereby repressing the glucose uptake system and inducing exopolysaccharide production, while a G-track sequences located in the first hairpin of RsaI bind to C-rich sequences of several sRNAs (Bronesky et al., 2019). In the related species Staphylococcus epidermidis, the sRNA RsaE is spatiotemporally expressed in biofilms. It exists in a full-length and a processed (20 nt shorter) form, and the two forms bind to the 5′-UTRs of different mRNAs: the full-length form facilitates extracellular DNA release, while the processed form regulates the central carbon metabolism (Schoenfelder et al., 2019).
In B. subtilis, the RsaE homolog RoxS is regulated by nitric oxide. It affects the ppnKB mRNA (encoding an NAD+/NADH kinase) by translation inhibition and destabilization. Interestingly, the latter is mediated by two independent mechanisms involving RNase III or RNase Y. Upon processing, RoxS also shows a change in the target specificity toward sucCD mRNA encoding succinyl-CoA synthase (Durand et al., 2015). The RoxS sRNA is regulated by an RNA sponge (Durand et al., 2021). Only few B. subtilis sRNAs were characterized functionally and available data were reviewed recently (Ul Haq et al., 2020). Similar to Staphylococcus, Bacillus also does not have ProQ but harbors Hfq, for which, however, no RNA chaperone properties were described yet (Zheng et al., 2016). Instead, CsrA was shown to promote the annealing of the sRNA SR1 with its ahrC mRNA target encoding an arginine repressor/activator protein (Müller et al., 2019). Furthermore, in Listeria monocytogenes, which together with B. subtilis and S. aureus belongs to Bacillales, the LhrC sRNAs destabilize the tcsA mRNA encoding a T-cell-stimulating antigen (Ross et al., 2019). Also in Listeria, the Rli27 sRNA is upregulated during the intracellular life cycle, when it targets a long 5′-variant to regulate a cell wall protein (Quereda et al., 2014). In contrast to the Bacillales members which have Hfq but not ProQ, Mycobacterium belongs to Actinobacteria and lacks both RNA chaperones (Olejniczak & Storz, 2017). In M. smegmatis, RNA chaperone-independent interactions were described between a C-rich loop of 6C sRNA and its multiple targets including dnaB mRNA, which is essential for replication (Mai et al., 2019).
3.4 Current and emerging sRNA topicsDespite the huge progress in knowledge on sRNAs in bacteria, certain topics still remain understudied and new aspects are emerging. As mentioned above, one of the current challenges is to identify the protein interaction partners of the sRNAs. Besides Hfq, the identification of ProQ as a global RNA chaperone (Smirnov et al., 2016) and of RapZ as an sRNA-specific adaptor protein that activates RNase E to degrade the GlmZ sRNA (Durica-Mitic et al., 2020; Göpel et al., 2013) demonstrated the importance of RBPs for RNA regulation. It is reasonable to propose that RBP binding to sRNA could be regulated by covalent modification of the RBP and/or the sRNA, or by their allosteric interactions with small molecule effectors, but such mechanisms were not shown experimentally yet. However, it was shown that interaction of the RBP RapZ with glucosamine-6-phosphate (GlcN6P) accounts for its second function in the GlmZ-glmS regulation paradigm: At low GlcN6P level, the GlcN6P-free form of RapZ stimulates the phosphorylation of a two-component system inducing the GlmY sRNA, which, in turn, sequesters RapZ to prevent GlmZ degradation. This leads to the accumulation of the base-pairing sRNA GlmZ, which activates the translation of glmS, the gene responsible for GlcN6P synthesis (Khan et al., 2020; Khan & Görke, 2020).
Another interesting topic is processed (truncated) sRNA forms. Shorter sRNA forms were often described, but for most of them, function is unknown. As mentioned above, sRNA processing could maturate/activate the sRNA or change its target specificity (Papenfort et al., 2009; Schoenfelder et al., 2019). Moreover, the processed sRNA can have a role in feedback regulation: Upon binding of the RBP RapZ to the GlmZ sRNA, the latter is cleaved by RNase E. Thereby, a processed sRNA form, GlmZ*, arises, which binds RapZ but does not allow for RNase E activation. The cleavage product GlmZ* competes with full-length GlmZ for RapZ binding, thus preventing complete GlmZ turnover and contributing to basal glmS expression (Durica-Mitic & Görke, 2019).
A continuous challenge is the identifications of sRNA targets and factors determining target specificity/prioritization. As mentioned above, high throughput methods such as MAPS, CLIP-seq, RIL-seq, and CLASH greatly contributed to the sRNA target detection in enterobacteria under specific conditions. In addition, bioinformatic prediction remains an important tool for target identification (Li, Edelmann, et al., 2021; Wright & Georg, 2018). Since most base-pairing sRNAs bind their targets by imperfect complementarity, theoretically many more mRNAs could be bound by an sRNA than experimentally found. Two studies addressed features distinguishing mRNA targets from nontargets of Hfq-dependent sRNAs in E. coli, showing the importance of the base-pairing extent, accessibility of the RNA seed regions necessary for the initial interaction, and no overlap between interaction site and Hfq binding site (Beisel et al., 2012; Bobrovskyy et al., 2019). Integration of such aspects in the bioinformatic predictions and combination of experimental and computational methods maximize target identification (Georg et al., 2020; Raden et al., 2020). It was proposed that subcellular RNA localization also influences target prioritization (Teimouri et al., 2017).
Subcellular sRNA localization in ribonucleoprotein (RNP) bodies organized by liquid–liquid phase separation (LLPS) is an emerging research topic (Irastortza-Olaziregi & Amster-Choder, 2021). The prokaryotic bacterial cell is spatiotemporally organized, having uneven distribution of macromolecules. It is known that in C. crescentus, tmRNA (responsible for trans-translation) is localized in a cell-cycle-dependent manner (Russell & Keiler, 2009). Furthermore, in E. coli and B. subtilis, the key endoribonucleases RNase E and RNase Y, which organize the RNA degradosome complexes in these bacteria, are localized at the cytoplasmic membrane, where they form short-lived foci (Hamouche et al., 2020; Strahl et al., 2015). Similarly, an RNase J-based degradosome forms foci at the cytoplasmic membrane in H. pylori (Tejada-Arranz et al.,
留言 (0)