
記住我





Since the revolutionary studies by Santiago Ramón y Cajal (1852–1934) and Camillo Golgi (1843–1926) [1, 2] – whose work is still a foundation of routine techniques of histopathologists and neuroscientists [3-5] – to modern developments of molecule-specific labeling and imaging in freely moving animals, microscopy-related technologies have constantly impacted upon the understanding of the healthy and pathological nervous system and made clear the importance of understanding the relationship between cell structure, localization and connections [3, 6, 7].
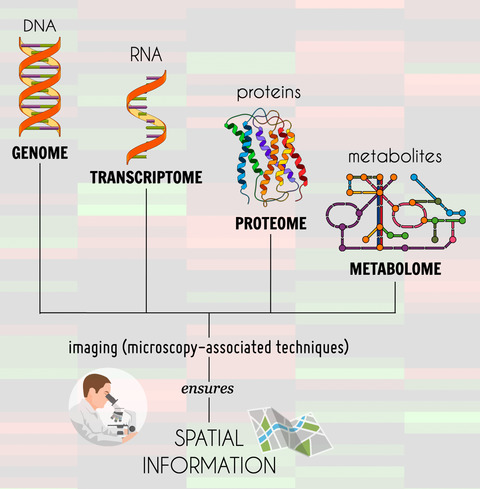
The collection of fields known as ‘omics’ also highlights the importance of connections. The omics fields, such as genomics, transcriptomics, proteomics and metabolomics, aim to identify and quantify a large number of elements associated with a physiological function and the connections between them. It has also led to ground-breaking findings, e.g., by identifying targets that could help to confirm diagnosis [8]. Omics approaches are particularly helpful when studying immunological functions such as neuroinflammation, as the techniques allow complex interactions to be drawn out. However, the processing of tissue removes the structural connections that are revealed by microscopy. In neuroinflammatory studies, changes in structural connections, cell morphology and localization of molecules are outcomes of interest, so a combination of both techniques is key. Several studies have integrated these two approaches to elucidate specific mechanisms, considering the correspondence between omics and microscopy.
In this review, we describe current applications of microscopy to image neuroinflammation, describing how changes in the shape and structure of specific cells can indicate neuroinflammatory processes. First, we will address some of the main neuroinflammatory responses and discuss the signs of neuroinflammation, focusing upon the involvement of the blood–brain barrier (BBB) and the phenotypical changes in glial cells, as well as the main markers that can be used to study neuroinflammation using microscopy techniques. Then, we discuss basic methodologies such as immunohistochemistry and immunofluorescence, which allow the microscopic observation of those neuroinflammatory markers. We will also emphasize the importance of two-photon microscopy to the in vivo real-time assessment of neuroinflammatory processes. Studies elucidating the function of the inflammatory components in healthy and disease conditions will be briefly mentioned as examples of the applications of these techniques. Lastly, we will discuss the newest omics techniques and how the molecular and imaging approaches may complement each other and be applied simultaneously to improve the comprehension of the mechanisms involved in neuroinflammation.
THE CONCEPT OF NEUROINFLAMMATIONThe term ‘neuroinflammation’ arose in the 1980s from observations using microscopy techniques which showed an accumulation of leukocytes around degenerating myelin and brain vessels in samples of multiple sclerosis (MS) [9]. Since then, the study of neuroinflammation has expanded greatly, with almost 5000 articles listed in NCBI PubMed for 2020 alone.
The process of neuroinflammation involves multiple types of cells and factors (Figure 1), which play different roles depending upon the context of the inflammation, duration, experimental model or disease, an understanding of which is complicated by inconsistency in terminology [10].

Neuroinflammation is characterized by several cellular and molecular processes leading to different cell phenotypes, mainly in astrocytes and microglia. These neuroinflammatory features (production and release of proinflammatory mediators and BBB leakage) can drive morphological changes in glial cells, which can be assessed through histological techniques with different markers (see the main text for details). BBB = blood–brain barrier
Neuroinflammation is mediated by signaling molecules known as cytokines, which are proteins produced and secreted by different cell types that mediate the immune and inflammatory responses [11]. However, it is worth mentioning that some of the ‘neuroinflammatory components’ are also involved in physiological function. For example, cytokines considered to be neuroinflammatory markers, such as interleukin (IL)-1, IL-18, IL-6 and tumor necrosis factor (TNF)-α, have already been described as key players in physiological mechanisms involved in memory consolidation [12]. Glial cells are also involved in mechanisms activated in non-pathological contexts. Microglia can be activated through IL-17 released from hippocampal neurons taking part in synaptic remodeling as part of memory consolidation [13]. The recent description of the importance of IL-17 from meningeal-resident γδ T cells for short-term memory [14] has also increasingly opened up the possibilities of studying ‘inflammatory’ cells and mediators in both health and disease.
Nevertheless, in pathological contexts, the inflammatory signaling that characterizes neuroinflammation might become detrimental and ultimately result in neuronal death [15, 16]. This process involves different cytokines and chemokines, as well as reactive oxygen species synthesis by microglia, astrocytes, infiltrated immune cells and endothelial cells. These mediators can potentially lead to harmful outcomes, such as edema, tissue damage and cell death [10, 11, 17, 18]. Glial cells orchestrate an inflammatory response in an extremely context-dependent manner, presenting different neuroinflammatory phenotypes based upon which pathways are activated, insult context and the degree of inflammation in experimental models, developmental stages and in health and disease conditions [10, 11, 19-22]. The situation is complicated somewhat by the lack of clear terminological definitions of individual cells in the CNS, such as terminology concerning astrocyte responses in physiological and pathological conditions [23]. Table 1 summarizes a variety of targets commonly used in light microscopy studies to describe different aspects of the neuroinflammatory process.
TABLE 1. Common targets and related outcomes regarding neuroinflammation markers Target Description Comments Histological-evaluated responses in different neurological conditions Damage to the blood–brain barrier (BBB) CLDN Claudin, a transmembrane protein family component of the tight junctions at the BBB In neuroinflammatory contexts, a BBB impairment may occur, weakening tight junctions and allowing peripheral molecules and cells to invade the CNS. In this sense, evaluating such proteins can be useful to relate its findings with other BBB integrity assays [39, 42, 178-180]CLD5 remodeling and changes in localization after stroke [39];
↑ microglia-associated CLDN5 in a model of SLE and after LPS stimuli [42];
↓ CLDN5 in a model of EAE [179];
↓ CLDN3 in MS [180]
OCLN Occludin, together with CLDN, is one of the main protein families that are part of the tight junctions of the BBB↓ in stroke [39];
↓ in models of inhaled cigarettes and e-cigarette [178]
ZO Zonula occludens, a family of intracellular proteins that provides anchoring sites for the tight junctions of the BBB ↓ in stroke [39] Microglia Iba-1 Ionized calcium-binding adaptor molecule 1 Expressed in myeloid cells. In the CNS is highly and selectively expressed by microglia and widely used to visualize the morphological spectrum between the classic M1 (amoeboid) and M2 (ramified) states. Also used to quantify soma size, perimeter, and to distinguish these two phenotypes (usually associated with another marker, e.g., CD68) [54, 103, 144, 175, 181-186]↑ 7 days after global ischemia [181];
↑ 24 h after LPS challenge [144];
↑ 24 h after HI [182];
↑ 3 days after TBI [54];
↑ after LPS challenge [183];
↓ in the white matter in DSCL [175]
CX3CR1 Fractalkine receptor or G-protein coupled receptor 13 (GPR13) A chemokine receptor involved in leukocyte adhesion and transmigration [59, 187] ↑ in the brainstem in hypertensive rats [59] P2RY12 Purinergic receptor or G-protein coupled receptor P2Y A purinergic receptor used as an indicator of ramified microglia (used as a marker of microglial branches) [188] Microglial P2RY12 deficiency ↓ neurotoxicity after OGD [188] OX-42 Cluster of differentiation 11b (CD11b), a surface protein from the integrin superfamily Expressed in myeloid cells (↑ in neutrophils and monocytes in the presence of pathogens). In the brain, it is a specific marker of microglia. Due to its rapid shift, CD11b could be considered an early marker of infection. It was blocked in experiments to evaluate microglial phagocytosis of newborn astrocytes [131, 189, 190] ↑ 8 h after reperfusion in ischemic stroke [131] CD45 Cluster of differentiation 45 Surface tyrosine phosphatase protein, a protein expressed in the membrane of hematopoietic cells. It is highly expressed in CNS-associated macrophages and expressed in low levels in microglia, allowing to differentiate macrophages from microglia. Expression levels may also indicate amoeboid or more branched morphology. CD45 is increased in disease-related conditions [64, 191-195]↑ around cortical plate lesions in a model of neonatal excitotoxic brain damage [191];
↑ after acute brain injury [192]
CD68 Cluster of differentiation 68 Transmembrane glycoprotein expressed in monocytes and in macrophages with phagocytic activity, usually suggesting microglia with ameboid morphology (M1 phenotype) [54, 175, 196].↑ after TBI [54];
involved in white matter damage [175];
↑ in morphologically activated microglia after cranial irradiation [196]
CD86 Cluster of differentiation 86 Microglia/ameboid morphology marker, a transmembrane glycoprotein constitutively expressed in immune cells. Usually, double staining for CD86 and Iba-1 is used to identify microglial phenotype [90, 197, 198]↑ in microglia after subarachnoid hemorrhage [197];
↑ after acute hypoxia [90];
↑ in a model of sepsis-associated encephalopathy [198]
CD206 Cluster of differentiation 206 Mannose receptor. Its expression provides anti-inflammatory and immunosuppressive responses of microglia, suggesting M2 phenotype polarization [62, 90, 197, 199-201]↑ after ischemia [62];
apparently not expressed in acute sepsis [201];
↑in TLR-4–/– mice after subarachnoid hemorrhage [197];
↓ after acute hypoxia [90]
Arg1 Arginase 1 Alternative phenotype marker of microglia, which expression might be up-regulated in the alternative activation phenotype [184] ↓ after ischemic stroke [184] Astrocytes GFAP Glial fibrillary acidic protein Intermediate filament protein of astrocytes. The cytoskeleton of reactive astrocytes may undergo hypertrophy and other structural changes, affecting GFAP expression [54, 119, 202, 203]↑ in obese rats after an HFD diet [119];
↑ expression in TBI and contusion trauma [202];
↑ after OGD [203]
S100B Calcium-binding protein Primarily produced by astrocytes and mainly expressed in the cytoplasm of these cells, but oligodendrocytes also express S100B. This protein is involved in calcium homeostasis, and as glial cells are sensitive to CNS disturbances, S100B is usually regarded as an indirect biomarker of neural injury [103, 204-207]↑ in a model of stroke-prone spontaneously hypertensive rats [204];
↓ in a model of diabetic ketoacidosis brain injury [205];
↑ in an in vitro model of MS [206]
Cytokines IL-1β Proinflammatory cytokine Inflammatory mediators usually produced by glial cells in neuroinflammatory events [59, 65, 162, 208, 209]↑ in hypertension [59];
↑ in MS [208]
IL-6 Usually considered as a proinflammatory cytokine, but may present pro- or anti-inflammatory roles depending on the context↑ in hypertension [59];
↑ in AD senile plaques [209];
↑ after stroke [162]
TNF-α Proinflammatory cytokine↑ in cerebral ischemia [65];
↑ after stroke [162]
Abbreviations: AD = Alzheimer’s disease; Arg1 = arginase 1; BBB = blood–brain barrier; CLDN = claudin; CNS = central nervous system; CX3CR1 = CX3C chemokine receptor 1; DSCL = deep subcortical lesions; EAE = experimental autoimmune encephalopathy; GFAP = glial fibrillary acidic protein; HFD = high-fat diet; HI = hypoxia–ischemia; Iba-1 = ionized calcium-binding adapter molecule 1; IL-1β = interleukin-1 beta; IL-6 = interleukin-6; KO = knock-out; LPS = lipopolysaccharide; MS = multiple sclerosis; OCLN = occludin; OGD = oxygen-glucose deprivation; OX-42 = anti-integrin αM, CD11b antibody; P2RY12 = purinergic receptor P2Y, G-protein-coupled receptor 12; S100B = S100 calcium-binding protein B; SLE = systemic lupus erythematosus; TBI = traumatic brain injury; TLR-4 = Toll-like receptor 4; TNF-α = tumor necrosis factor-alpha; ZO = zonula occludens. WHAT TO OBSERVE: THE SIGNALS OF NEUROINFLAMMATIONIn this section, we introduce the cell markers used to study BBB damage in neuroinflammatory contexts and the brain clearance ‘glymphatic’ system. We also discuss the cells involved in the neuroinflammatory responses, together with the immunological synapse that comprises the interaction between nervous and immune cells. These are the main targets to be observed when using a microscope as a tool to study neuroinflammation.
Damage to the BBBAlthough seen as a structure separating the nervous system from the periphery, the BBB also regulates the passage of substances into and out of the nervous system, and damage to the BBB is a key point in several inflammatory processes.
The BBB is essential to maintain the microenvironment of the CNS distinct from the periphery, regulating the entry and exit of solutes and the entry of peripheral leukocytes [24]. Its function is performed by specialized blood vessel endothelial cells, which have limited vesicular transport and are connected through highly specialized tight junctions, preventing paracellular transport [25, 26]. The tight junctions are formed by the claudins (CLDN), occludin (OCLN) and the zonula occludens protein family (ZO-1, -2 and -3) [27]. Pericytes and astrocytes are cellular components in close contact with the endothelium, influencing and participating in the complex maintenance of the BBB [26, 28].
The BBB is disrupted in several neuroinflammatory contexts [29] such as cerebral ischemia [30, 31], Alzheimer’s disease (AD) [32, 33] and MS [34], and is also affected by systemic inflammation, e.g., induced by systemic injections of lipopolysaccharide (LPS) [35]. Loss of BBB integrity can also affect neurodevelopment, as shown in a model of prenatal exposure to valproic acid in rats [36] and neonatal hypoxic–ischemic events [37]. Sophisticated imaging techniques such as computerized tomography (CT), magnetic resonance imaging (MRI) and near-infrared fluorescence (NIRF) are widely used in these studies, but the limitation of analysis to macroscopic scales places microscopy as an important tool to elucidate the molecular mechanisms involved in the neuroinflammatory process [27].
Immunohistochemistry (IHC) has been used in brain samples from both epileptic patients and rats to detect increased anti-albumin labeling in the hippocampus [38]. Because albumin is not expected to cross the BBB under normal conditions, this suggests a loss of barrier integrity [24]. The paper by van Vliet et al. [38] demonstrated similar results in chronically epileptic rats using immunofluorescence (IF) techniques. This finding was subsequently confirmed by injecting fluorescein dye into the tail vein of the animals and visualizing the infiltrated dye with a confocal microscope. As fluorescein does not penetrate the CNS under normal conditions and can bind to albumin, it offers a confirmatory method to analyze the infiltration. The albumin signal was also co-labeled with astrocyte, neuronal and microglial markers to demonstrate the relationship of these cells with invading molecules [38].
In addition to analyzing the leakage of some substances through the BBB, targeting junction proteins can provide relevant data on BBB integrity. Decreased expression of junction proteins CLDN-5, OCLN, ZO-1, α-catenin and vascular endothelial cadherin was detected by IF in pre-reactive inflammatory lesions from MS human brain samples [34]. One-time and repeated imaging were performed in a mouse model of ischemic stroke with the CLDN-5 gene coupled with the green fluorescent protein (GFP) reporter gene, in which green fluorescence was detected using two-photon microscopy, thus providing evidence of dynamic changes in tight junctions in this pre-clinical ischemic stroke model [39]. Also, using GFP expression under the control of CLDN-5 promoter, two-photon microscopy revealed remodeling of the tight junctions preceding the invasion of a peripherally injected dye that normally does not cross the BBB in an experimental autoimmune encephalitis (EAE) model. Interestingly, this dynamic remodeling of the tight junctions precedes the EAE onset [40].
In a blast-induced traumatic brain injury (TBI) model, a decrease in the fluorescence intensity of ZO-1, CLDN-5 and OCLN suggested a BBB leakage that was confirmed by analyzing the fluorescent tetramethylrhodamine isothiocyanate–dextran (TRITC-dextran) dye invasion into the brain [41]. An interesting method to quantitatively analyze the damage to the BBB is the use of dyes of different sizes. In a model of sustained inflammation induced by systemic injections of LPS, only dextran-Texas red (10 kDa) crossed the BBB, while dextran-tetramethylrhodamine (40 kDa) and dextran–fluorescein (70 kDa) did not [42]. Thus, as described so far, microscopy can assist in fundamental issues related to BBB integrity from differences in immunoreactivity and location of junction proteins involved in BBB integrity to the degree of barrier breakdown assessed through fluorescent dyes.
IHC and IF allow the co-localization of inflammatory cytokines with different cell markers to determine the involvement of specific cell types in the neuroinflammatory process. Four hours after systemic LPS administration, stromal cell niches in leptomeningeal and choroid plexus and epithelial cells from the choroid plexus and hippocampal vessels were the main producers of chemokine (C-C motif) ligand 2 (CCL2), chemokine (C-X-C motif) ligand 1 (CXCL1), CXCL2 and IL-6. Interestingly, after 24 h the main source of mediators, such as CCL11 and CXCL10, were astrocytes, evidenced by co-labeling with glial fibrillary acidic protein (GFAP). The use of histological techniques also showed that the expression of cytokine receptors following systemic LPS administration occurred specifically in the astrocytic end-feet, a part of the cells that is in contact with capillaries and which plays a role in BBB maintenance [43].
Visualizing fluorescent tracers in the cerebral cortex using two-photon microscopy allowed the elucidation of a brain waste clearance system: a perivascular pathway that enables the transport of cerebrospinal fluid (CSF) into and out of the brain parenchyma. The same study showed that aquaporin 4 (AQP4) in astrocyte foot processes is fundamental to fluid dynamics [44]. The glial-based clearance system was termed the ‘glymphatic system’. The same research group later used in vivo two-photon and ex vivo fluorescence microscopy to find that cerebral arterial pulsatility is also critical to CSF transport into and through the brain [45]. Also, light-sheet fluorescence microscopy has been applied as a new way to study the glymphatic system, reducing some of the limitations such as the need for antibodies, due to the use of CSF tracers (for details, see Bèchet et al. [46]). The wide range of approaches in these studies demonstrates that microscopy is a fundamental tool for describing this important clearance system in the brain.
Cell types involved in the neuroinflammatory responseGlial cells – mainly microglia and astrocytes – and invading leukocytes orchestrate the neuroinflammatory response following BBB disruptions. Microglia cells are the CNS macrophages and assume different phenotypes within a spectrum ranging from M1 (proinflammatory phenotype) to M2 states (anti-inflammatory phenotype) [47, 48]. The M1 phenotype is characterized by secretion of proinflammatory cytokines such as TNF-α, IFN-γ and IL-6 [49], whereas the M2 phenotype is related to anti-inflammatory cytokines such as IL-4 and IL-10 [49]. As well as differences in the cytokines released by each microglial phenotype, the shape of the microglia themselves following stimulation also varies and the different shapes reflect their differing function. For example, following LPS challenge, M1 microglia show a profile with an ameboid shape [50], aiding its ability to phagocytose pathogens in the CNS. In contrast, M2 morphology presents ramified branches that enhance its surveillance role [51]. It is important to point out here that over-simplifications of these phenotypes should be avoided when describing not only the state of microglia, but also the outcomes of a particular neuroinflammatory event [52].
Microglia polarization analyzed by immunostaining and further confirmed by flow cytometry has been employed in middle cerebral artery occlusion (MCAO) models [53]. Flow cytometry has been employed to confirm the inflammatory response and microglial polarization using F4/80 labeling, which is a glycoprotein found on the surface of macrophages and resting microglia [53].
Usually, microglia alterations are widely used to suggest a neuroinflammation condition in the CNS using the ionized calcium-binding adaptor molecule 1 (Iba-1) marker [54-56]. Some authors applied double-labeling of the Iba-1 as a marker for all microglial cells, labeling the soma, with LN3 (a monoclonal antibody used as a microglial activation marker as it recognizes the major histocompatibility complex (MHC)-II), to evaluate microglial hypertrophic and ameboid morphology [57]. Another interesting marker, P2Y purinoceptor 12 (P2RY12), was double-stained with Iba-1 to enhance the labeling of the microglial ramifications [57, 58]. Also, CX3C chemokine receptor 1 (CX3CR1) fractalkine receptor is expressed on the surface of macrophages and microglial cells and responds to CX3CL1 (fractalkine) [59-61], and is considered a potential marker for macrophages and microglial cells.
Although it can lead to confusion with neutrophils, CD11b is used as a marker for microglia, as these cells express this protein [56, 62]. Conversely, myeloperoxidase protein (MPO) can be used as a marker for neutrophils [63], allowing discrimination of neutrophils from microglia. As mentioned above, Iba-1 is widely considered a microglia marker; however, this antigen does not differentiate microglia from monocytes. CD45 could help in this case as it is more expressed in monocytes than in microglia, so both Iba-1- and CD45-positive cells – particularly round-shaped cells – are probably monocytes, while Iba-1-positive and CD45-negative cells with processes are microglia [62, 64]. Another helpful marker is CD68 (a macrophage/phagocytic activity marker) which appears after ischemia in the border zone of the lesion, but invades the ischemia core 7 days after the insult [56, 62].
The microglial inhibitor AZD8797 (a CX3CR1 inhibitor) was used in a study designed to observe microglial-dependent neuroinflammatory responses. A significant increase in the percentage of IL-1β+ and IL-6+ cells (observed by IHC) caused by hypertension was mitigated by blocking microglia with AZD8797 [59]. In a model of brain ischemia, immunofluorescent staining was used to co-label cells with anti-Iba-1 and anti-TNF-α, demonstrating that microglia were the main cell type responsible for the TNF-α increase observed in the brain tissue. As IF was performed after 12 h and 1, 3 and 7 days following ischemia induction, it allowed a sequential analysis of the role of microglia in establishing the neuroinflammatory process [65].
Due to the extensive range of possible microglial phenotypes, double-labeling may differentiate the pattern of the response of these cells in a particular situation. Moreover, studies should consider not only the number or optical density of cells immunoreactive to these markers, but also cell morphology and time–course of the event. These factors further reinforce the need to use microscopy techniques to study cellular responses in neuroinflammation [64, 66].
Astrocytes also play an important role in different CNS diseases [67] such as stroke [68], TBI [69], AD [70-72], amyotrophic lateral sclerosis (ALS) [73] and other neurodegenerative diseases [67, 72, 74]. Astrocytes and microglia act in a coordinated manner to maintain homeostasis [75], influencing each other in the pathophysiology and inflammatory responses [76-
留言 (0)