
記住我

Recruitment of new hippocampal neurons is part of memory encoding where active neuronal ensembles form contextual representations of discrete experiential episodes. Tasks involving retrieval require reactivation of the representations formed during encoding (Guzowski et al., 1999; Chawla et al., 2005; Garner et al., 2012; Pevzner et al., 2012; Josselyn et al., 2015; Tonegawa et al., 2015a,2015b; Eichenbaum, 2016). If those representations remapped (ie, a new cellular ensemble was recruited, rather than reactivation of the cells comprising the previously formed representation) this should theoretically result in a retrieval error. Therefore, switching the memory system back to a state of encoding would prove maladaptive in situations where retrieval is necessary to perform a memory task unless new information was at hand. In this case, it would be adaptive to incorporate that new information into the memory trace and theoretically, retrieval and encoding would occur together to update that representation.
Furthermore, it is hypothesized that posttraumatic stress disorder (PTSD) involves impairments in memory-updating mechanisms where incorporation of new information (eg, safety signals) is not encoded at a functional level which may reflect an inability to remap hippocampal contextual representations (ie, trauma-related representations are reactivated rather than incorporating safety signals into existing memory traces) (Maren et al., 2013; Morrison and Ressler, 2014; Giustino et al., 2016; Liberzon and Abelson, 2016; Elsey and Kindt, 2017; Lee et al., 2017; Sheynin and Liberzon, 2017). The pathophysiology of anxiety disorders such as PTSD is characterized by noradrenergic dysregulation (Hendrickson and Raskind, 2016). We believe the locus coeruleus (LC), the site of noradrenergic cell bodies, plays a role in memory updating, specifically by biasing the system towards encoding. This suggests that individuals with PTSD may be experiencing an inability to engage this transition to encoding through dysfunction of the LC norepinephrine (NE) system.
The LC responds to salience cues including novelty and sends a major noradrenergic projection to the dentate gyrus (DG) (Aston-Jones and Bloom, 1981; Vankov et al., 1995; Berridge and Waterhouse, 2003; Harley, 2007; Aston-Jones and Waterhouse, 2016). LC activation causes NE release (Blackstad et al., 1967; Fuxe et al., 1968; Ungerstedt, 1971; Lindvall and Björklund, 1974; Pickel et al., 1974; Ross and Reis, 1974; Babstock and Harley, 1992; Frizzell and Harley, 1994) and induces downstream hippocampal plasticity (Bliss et al., 1983; Neuman and Harley, 1983; Stanton and Sarvey, 1985; Lacaille and Harley, 1985; Gray and Johnston, 1987; Hopkins and Johnston, 1988; Harley, 1991; Klukowski and Harley, 1994; Walling and Harley, 2004; Almaguer-Melian et al., 2005; Lashgari et al., 2008; Lemon et al., 2009; Lim et al., 2010; Walling et al., 2011; Hagena et al., 2016), effects which are β-adrenergic receptor (BAR)-dependent (Kitchigina et al., 1997). It is thought that LC-NE activation induces changes in network dynamics occurring at critical times when learning is necessary to promote adaptive behavior, for example, reversal learning (Sara et al., 1994). These configurations function to reset the system (Bouret and Sara, 2005) as novelty-associated activation of the LC causes the hippocampus (HPC) to recruit a new population of neurons to represent the immediate context (ie, global remapping) (Grella et al., 2019). This observation is consistent with the idea that NE provides a reset signal promoting global remapping in the HPC in the presence of new information and our hypothesis that it can bias memory towards encoding. This hypothesis suggests that the effect of modulating NE on memory will depend on the stage of training.
Although NE has been shown to mediate different stages of memory (McGaugh et al., 1990; Do Monte et al., 2008) it is unclear how it is involved in updating memory. To assess this, we investigated how activation and blockade of BARs exert modulatory influence during learning and recall. We tested whether infusions of the BAR-agonist isoproterenol (ISO) would impair working and reference memory retrieval by switching the system to encoding when it is potentially maladaptive (eg, when retrieval of a previously established ensemble is required to complete a task). Given that LC neurons exhibit plasticity as a function of environmental contingency changes to promote adaptive behavior (Sara and Segal, 1991), we also tested whether ISO would, in contrast, enhance cognitive flexibility by promoting encoding when it is potentially adaptive (eg, when new information is presented) and whether these effects could be blocked with the BAR-antagonist propranolol (PRO).
Materials and methods SubjectsEighty-seven adult male Fischer-344 rats were purchased from Harlan (Indianapolis, Indiana, USA) at 16 weeks old (~350–375 g) for experiment 1 & 2 and at 10 weeks old (~300–325 g) for experiment 3. The number of rats included in each experiment and group are listed in Table S1, Supplemental digital content 1, http://links.lww.com/BPHARM/A69 and in the figure legends. Rats were housed in standard transparent Plexiglas cages (47.6 cm L × 26.0 cm W × 20.3 cm H), pair-housed initially and then single-housed after surgery. They were kept on a 12:12 h reverse light cycle (lights ON at 7 p.m.) and provided with food (Teklad lab diet, Envigo) and (tap) water ad libitum until they recovered from surgery after which the animals in experiment 1 were food restricted to 90% of their free-fed weight. Animals in experiments 2 & 3 remained on an ad libitum diet. All procedures were approved by the Wilfrid Laurier University Animal Care Committee in accordance with the guidelines of the Canadian Council on Animal Care.
SurgeryFor 4 consecutive days prior to surgery animals were weighed, handled for 15 min and given 20 g of a nutritionally complete dietary supplement containing trimethoprim/sulfamethoxazole antibiotic (MediGel TMS; ClearH20, Westbrook, Maine, USA) in addition to their regular diet in their home cage. The following day, rats underwent implantation of a bilateral guide cannula. Several days prior to surgery, two 22-gauge stainless steel guide cannulas (Plastics One, Roanoke, Virginia, USA) were cemented together to form a bilateral cannula and left to dry. The next day they were autoclaved and again left to dry. At the start of surgery, rats were deeply anesthetized with 5% isoflurane and 70% oxygen (induction) and maintained at a level of 2–3% isoflurane for the duration of the surgery. They were anchored in a stereotaxic frame with ear bars to ensure a flat skull surface and prepped for aseptic surgery. Rats were administered a subcutaneous (s.c.) injection of ketoprofen (Anafen; Sigma Aldrich, Oakville, Ontario, USA; 0.15 ml of a 10 mg/ml solution) for general analgesia, and 3 ml of sterile physiological saline (0.9% NaCl; s.c.) for fluid replacement in case of blood loss. A midline incision was made on the scalp and six holes were drilled. Each rat was implanted with the bilateral cannula (8 mm in length, Plastics One) aimed at the dorsal DG with the coordinates: AP −3.3 mm, ML ±2.1 mm, DV −4.2 mm (from skull) relative to Bregma (Paxinos and Watson, 2013). Cannulae were anchored to the skull with four skull screws (#0-80, Plastics One) and dental acrylic. At the end of surgery, stainless steel stylets (flush with guide) were screwed into the cannulae to ensure patency and rats were placed on a heating pad for 1 h. They were given an additional 0.15 ml injection (s.c.) of ketoprofen (Sigma Aldrich) 24 h later and allowed 7 days for recovery undisturbed except for daily weighing. During the first 4 days of recovery, rats continued to receive 20 g of TMS in their home cage and were given their regular diet mixed with water in a mashed form in addition to regular chow pellets.
Drugs and infusionsRats received either (−)-isoproterenol bitartrate [ISO; 10 µg/µl (5 µg/side) dissolved in sterile saline; Sigma Aldrich] or (±)-propranolol hydrochloride (DL) [PRO; 3 µg/µl (2.5 µg/side) dissolved in sterile saline; Sigma Aldrich]. Given that few experiments have targeted the DG with these specific drugs in awake, freely moving animals, the doses we chose were on the basis of a literature search (Table S2, Supplemental digital content 1, http://links.lww.com/BPHARM/A69) (Alsene et al., 2011; Cahill and Alkire 2003; Garrido Zinn et al., 2016; Hansen and Manahan-Vaughan 2015; Lethbridge et al., 2104; Liu et al., 2012, 2014; Ramirez et al., 2013; Rowland et al., 2011; Straube et al., 2013; Sun et al., 2005; van Stegeren, 2008). We decided to infuse 5 µg in the DG of each hemisphere since Geyer and Masten (1989) found that infusion of a similar amount resulted in an increase in diversive (reconnaissance-like) exploration. For PRO, no studies had targeted the DG specifically. Ji et al. (2003) and Chai et al. (2014) found impaired memory consolidation and a blockade of NE-facilitated memory enhancements when they targeted the CA1 (5 µg per side). However, Hatfield and McGaugh (1999) and Barsegyan et al. (2014) found spatial memory impairments and diminished NE-facilitated memory enhancements with smaller amounts (0.3 and 1 µg) when targeting the BLA. Therefore, we decided to use 1.5 μg for PRO. For each infusion, stylets were unscrewed from each rat’s cannulae and a 30-gauge infusion cannula (1 mm below pedestal) connected via polyethylene tubing (PE-10) to a 10-μl Hamilton syringe mounted onto a microfluidic infusion pump (Harvard Apparatus, model: 70-2000) was inserted into the guide cannulae. Rats were infused with 0.5 μl on either side of the brain at a rate of 0.5 μl/min and the infusion cannula left in place for 1 min postinfusion to ensure the liquid had diffused from the injection site.
Experiment 1: delayed nonmatch to position (DNMP) ApparatusWe used a radial arm maze (122 cm in diameter; Stoelting Co., Wood Dale, Illinois, USA), which consisted of 12 gray, equidistantly spaced, polyethylene arms (50 cm L × 10 cm W × 13 cm H) that radiated from a small circular rotating central platform. The maze rested on a table, elevated 84 cm from the ground, located in the center of the room (2.44 m L × 2.24 m W × 2.95 m H) and extravisual cues (geometric shapes) were positioned on the walls. Other visual cues included a computer in one corner of the room.
Pilot experimentTo decide which arm separations to use for the task, we ran a pilot study with animals that did not undergo surgery. These rats received habituation and pretraining trials and then six acquisition-training sessions (see below). Each acquisition-training session consisted of six trials (sample + choice) per day to assess performance on arm separations 1–6 (order counterbalanced). We measured latency to obtain the reward, the number of errors made and the percentage of trials where a correct response was made for the last 3 days.
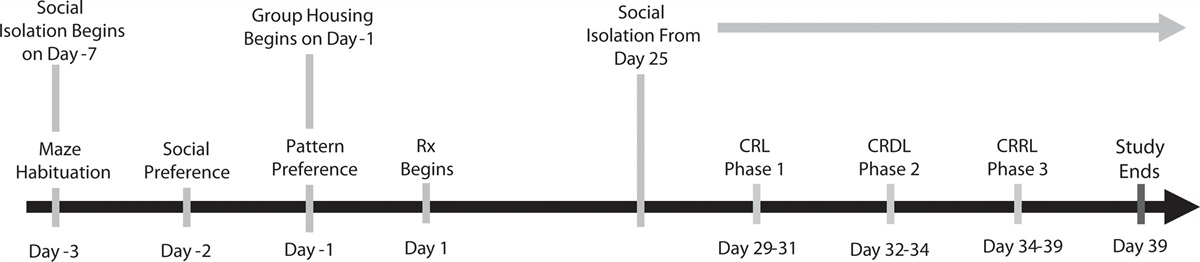
ProcedureThe DNMP task consisted of 5 stages: (1) habituation, (2) pretraining, (3) acquisition, (4) testing and (5) curtain probe test. Timeline and task schematic outlined in Fig. 1a–c.
 Fig. 1:
Fig. 1: (a) Experiment 1 timeline. (b) Timeline for infusions on test days. (c) Schematic showing DNMP task. During the sample phase, one arm is open and baited and animals are trained to obtain a reward from this arm. After a delay of 10 min, in the choice phase, animals are placed back in the maze and presented with a choice between the previously rewarded arm and the new arm. The sample and choice arms are separated by either 2 (S2) or 5 (S5) arms making the task DG-dependent or DG-independent respectively. (D-I) test day - animals were tested on four conditions on 4 different days with each test day separated by a washout day. Rats were assigned to either the ISO (blue; n = 20) or PRO (red; n = 20) group and received a drug infusion either 3 min prior to the sample phase (PS) or the choice phase (PC) and were tested in the S2 (checkered) or S5 (striped) condition. TRIAL 1: habituation trial (not shown); TRIAL 2: baseline trial (BASE) - animals infused with saline; TRIAL 3: test trial (TEST) - animals received drug treatment they were assigned to. Significant differences denoted with asterisks *P < 0.05, **P < 0.01, ***P < 0.001. DNMP, delayed nonmatch to position; DG, dentate gyrus; HC, home cage; ISO, isoproterenol; PRO, propranolol; S2, two arm separation; S5, 5 arm separation; PS, pre-sample; PC, pre-choice.
Habituation and pretrainingHabituation lasted 4 days. On day 1, rats were given one 20-min habituation trial where they freely explored the maze. All 12 arms were open and baited with a reward placed in a small plastic gray cup at the end of the arm. The next day, rats were given two 10 min trials (intertrial interval = 1 h) in the maze with six arms open and baited. For the next 2 days, they were given two 5 min trials a day with three arms open and baited. On day 5 rats began pretraining. Pretraining lasted 10 days. During this phase, rats were given two trials/day with only one arm open and baited. The goal was to train the rats to retrieve the reward in less than 2 min. By the 10th day, all rats could do this. On day 14, rats began acquisition training.
Acquisition trainingAnimals received 4 trials/day. Each trial consisted of two phases: sample and choice, separated by a 10-min delay. During the sample, all arms except the sample arm were blocked off. The rat was placed in the center of the maze and permitted to visit the sample arm and obtain half a Fruit Loop reward. Latency to obtain the reward was recorded. Once the animal retrieved the reward, he was left in the maze an additional 10 s to promote memory for the sample arm location. The rat was then placed back in his home cage and 10 min later tested on the choice phase. During the 10 min delay, the maze was rotated, preserving arm location, yet eliminating the possibility of odor being used as an intramaze cue. During the choice phase, the previously rewarded sample arm was now unrewarded. An additional correct arm was open and rewarded. Correct arms varied in distance from the sample arm by spatial separation of 2 (S2), or 5 (S5) arms (Fig. 1b,c). Cups in each of the two arms appeared identical from afar and both contained half a Froot Loop, but the cup in the unrewarded arm had a mesh overlay preventing access to the reward. Latency to reach the reward, choice accuracy and number of errors were recorded. When the incorrect arm was chosen, the rat was permitted to self-correct. Re-entry into the incorrect arm was considered an additional error. When a correct choice was made, an additional full Froot Loop was given in the home cage immediately after the trial ended. Rats were given four trials (sample + choice) per day (2× S2 & 2× S5) of pseudo-randomly presented combinations of sample + correct arms (Tables S3 & S4) [intertrial interval (ITI) = 90 min until a criterion of 4/6 correct choices were made on S5 trials across 3 consecutive days]. Criterion was reached within 6–7 days. One hour after the last acquisition training trial, stylets were unscrewed from each rat’s cannulae, and the infusion cannula was inserted to make sure the cannula was not blocked. The infuser was left in the cannula for 2 min on each side of the brain to simulate what would occur during testing, but no fluid was delivered. This was done in attempt to reduce the elicitation of nonspecific stress response on test day. Following this, the dust caps were screwed back in and animals were then returned to their home cage.
Test dayUsing a balanced Latin Square design (Table S5, Supplemental digital content 1, http://links.lww.com/BPHARM/A69), animals were tested on 4 conditions on 4 different days with each test day separated by a one-day washout (Fig. 1a). We used a 2 × 2 × 2 design with a between-subject factor of GROUP (drug treatment), a within-subject factor of INFUSION TIME, and a within-subject factor of ARM-SEPARATION. Rats were assigned to a drug treatment following acquisition (ISO or PRO). This remained constant throughout testing. On test day, rats were infused 3 min prior to either the sample phase (pre-sample) or the choice phase (pre-choice) and were tested on S2 and S5 conditions. So, all animals were tested on pre-sample-S2, pre-sample-S5, pre-choice-S2 and pre-choice-S5. On each of the 4 test days, instead of receiving two S2 and two S5 trials (as in training), animals received all four trials in the condition they were being tested (all S2 or all S5) allowing TRIAL to be included as an additional within-subject factor making a 2 × 2 × 2 × 3 design. Trial 1: Habituation - like the previous day, infusion cannulae were inserted but no fluid was infused; Trial 2: Baseline - animals were infused with sterile saline (0.9% NaCl); Trial 3: Test - animals received either ISO or PRO. In our analyses, we compared baseline to test (see Table S6, Supplemental digital content 1, http://links.lww.com/BPHARM/A69 for all conditions). During testing, the latency to obtain the reward, the number of errors, and the percent correct trials were measured. The next day after the final test day, rats were given one more washout and the following day, a curtain probe test.
Washout sessions and curtain probeBetween each test day, animals were given a washout day that was identical to acquisition training to allow the drug to clear before recommencing testing. To ensure that animals were using extramaze cues rather than intramaze or interoceptive cues to complete the DNMP task, following the last washout day animals were given a curtain probe. The procedure for both the washout sessions and the curtain probe was identical to acquisition training. For the curtain probe, the exception was that a blue curtain was hung from the ceiling in a circular fashion, surrounding the maze such that animals could not see any of the cues in the room except for the webcam above and a partial view of a few ceiling tiles.
Experiment 2: elevated plus maze ApparatusTo assess the effects of ISO and PRO on locomotion and anxiety, a separate group of rats were tested using a gray polyethylene elevated plus maze (EPM) (Stoelting Co., Wood Dale, Illinois, USA) consisting of two open and two closed runways (50 cm L × 10 cm W × 40 cm H) elevated 40 cm from the ground. EPM testing took place in a smaller room (1.83 m L × 1.78 m W × 2.95 m H) where the maze was positioned in the center. A separate group of animals was used since there were no drug-naïve animals in the DNMP experiment to serve as the vehicle group for EPM testing.
ProcedureRats underwent similar handling and surgical procedures as above. Following recovery, rats were split into three groups: ISO, PRO and vehicle. Using the same doses as above, rats were given a bilateral intra-DG infusion of either ISO, PRO, or vehicle and then 3 min later tested in the EPM. Rats were placed at the junction of the four arms at the beginning of the session. Their behavior was monitored for 5 min. Anxiety-like behavior was assessed by measuring the percentage of time spent in the open arms of the maze compared to the closed arms, and the number of entries into the open and closed arms. General locomotor activity was assessed by measuring total number of arm entries, total distance travelled, number of line crossings, mean speed, and the time spent immobile.
Experiment 3: Barnes maze ApparatusThe Barnes maze (Barnes, 1979) consisted of a gray circular polyethylene disk (122 cm diameter) with 20 circular equidistant holes (10 cm diameter; 9.65 cm between holes) located around the perimeter of the maze (1.3 cm from the edge). The maze was elevated 90 cm from the ground and beneath each hole was a slot where an escape box (35.56 L × 13.34 cm W × 10.16 cm H) or a ‘false’ escape box (11.43 cm L × 13.34 cm W × 1.9 cm H) could be inserted. For any given trial 19/20 holes were connected to a false escape box and only one hole lead to the true escape box. The false escape boxes were significantly smaller than the true escape box; therefore, rats could not escape the maze via these boxes. Their main purpose was to conceal any visual cues that may be apparent from a distance or through an open hole. Four bright white lights (150 W) were mounted above the maze, which illuminated the entire maze area. The rest of the room was dark when testing. Animals were motivated to escape from the brightly lit, open platform into the dark, recessed escape box due to their natural tendency to seek out dark, closed spaces. The maze was in the center of a large room (4.5 m L × 3.35 m W × 2.95 m H) and extra-maze visual cues (geometric shapes) were positioned on the walls. Other visual cues included several desks and cabinets.
ProcedureThe experiment consisted of nine distinct phases: (1) habituation (2) acquisition training (3) acquisition probe test (4) retraining (5) curtain probe test (6) second retraining (7) reversal training day 1 (8) reversal training days 2–5 and (9) reversal probe test (Exp. timeline and task schematic Fig. 3a,b).
 Fig. 2:
Fig. 2: Rats were tested in the EPM to determine if either ISO or PRO affected locomotor behavior. We measured (a) total distance traveled (b) mean speed (c) number of line crossings and (d) time spent immobile and found no effect of either VEH (gray; n = 5), ISO (blue; n = 5) or PRO (red; n = 5). Rats were also tested in the EPM to determine if either ISO or PRO affected anxiety-like behavior. We measured (e) the number of arm entries into the open and closed arms. We also measured (f) percentage of time the animals spent in the open arms, closed arms, and the start area. We found no group differences and no effect of ISO or PRO on anxiety-like behavior. All animals spent more time in the open arms. Significant differences denoted with asterisks *P < 0.05, **P < 0.01, ***P < 0.001. EPM, elevated plus maze; ISO, isoproterenol; PRO, propranolol; VEH, vehicle (saline).
HabituationRats were given one 5 min habituation trial where they freely explored the maze and could descend into the escape box. Once they entered the escape box, they were permitted to stay in the box for 30 s and were then removed and placed back into the center of the maze until the end of the 5 min period.
Acquisition trainingEach trial (except habituation) began with a 5 s acclimatization period during which the rat was being held in the start box in the center of the maze. Trials began automatically after the 5 s delay and the start box was lifted. For the first 4 days (A1–A4), rats were given three trials/day and during the following 8 days (A5–A12) this was reduced to two trials/day for a total of 28 trials with an intertrial-interval (ITI) of 2 h on all days. Including the habituation, trial animals received a total of 29 trials prior to the Acquisition Probe Test. Animals learned the spatial location of the escape box, which stayed consistent. Each trial lasted up to 5 min during, which, ANY-maze software recorded the latency to reach the escape hole, total distance traveled and path efficiency (see below for description). The experimenter recorded the number of reference errors the animal made prior to reaching the escape hole, the number of hole deviations there were between the first hole visited and the escape hole, and the search strategy used to find the escape hole. If the rat did not find the escape hole in the time allotted, it was gently guided to the escape box. Once the rat was inside, it remained there for 30 s before returning to its home cage. For all training trials, rats were grouped into squads of 3–4 where all members of a squad completed a given trial before subsequent trials were run.
Cardinal direction at startBefore testing began, rats were pseudo-randomly assigned to one of four possible escape locations. These locations were equidistantly positioned at 90-degree intervals (north, west, south, east). This was to prevent odor cues from becoming saturated around any one hole, although the maze was cleaned with 70% ethanol between trials to eliminate any odors. Given that each rat was placed in a holding box for a 5 s acclimatization period at the start of each trial, we could not choose the direction the rat would be facing when the trial began. To ensure that this was counterbalanced for north, west, south and east directions, the videos were scored (n = 850) by a researcher blind to the conditions of the experiment. The results are listed in Table S7, Supplemental digital content 1, http://links.lww.com/BPHARM/A69. This was necessary to assess whether rats were using a fixed motor response to find the escape hole.
MeasuresTo record behavior in all three testing rooms, a webcam connected to a computer running ANY-maze tracking software (Stoelting Co.) was mounted above each apparatus on the ceiling and behavior was tracked using ANY-maze software (Stoelting Co.).
Path efficiency is represented as an index of the efficiency of the path taken by the rat to get from the first position in the test (start) to the last position (escape hole). A value of 1 is indicative of perfect efficiency (eg, the animal moved in a straight line from the start to the escape hole). It is calculated by dividing the straight-line distance between the first and the last position by the total distance traveled by the rat. This measure was not used during probe sessions, as it cannot be analyzed across time.
Reference errors were recorded as a rat dipping its head into any hole other than the escape hole. Repeated dips into the same hole were considered a single error.
Hole deviations were quantified as the number of escape holes between the true escape hole and the location in which the animal’s head, first entered a false escape hole. This ranged between 0–10.
There were three possible search strategies: (1) random (RD) – this occurred when the animal moved about the maze in a random, unsystematic manner, searching the same hole more than once and moving into the center of the maze often. (2) Serial (SE) – animals that used a serial search strategy first visited a hole more than two-hole deviations away from the escape hole and then in a serial fashion systematically checked adjacent holes until reaching the escape hole. The animals' search path was classified as serial even if he did not make any errors but visited a location at the edge of the maze more than two holes away. (3) Spatial (SP) - this search strategy occurred when a rat moved directly from the center of the maze to the correct escape hole or any hole within two-hole deviations away on either the left or right side of the escape hole.
One hour after the last acquisition training trial, stylets were unscrewed from each rat’s cannulae, and the infusion cannula was inserted to make sure the cannula was not blocked. The infuser was left in the cannula for 2 min on each side of the brain to simulate what would occur during testing, but no fluid was delivered. This was done in attempts to reduce the elicitation of nonspecific stress response during the acquisition probe test. Following this, the stylets were screwed back in, and the animals were returned to their home cage.
Acquisition probeRats were given a 5 min acquisition probe where the escape box was removed and replaced with a false escape box. The maze was rotated to ensure the animals were using extra-maze visuospatial cues to find the escape hole. The maze was divided into 20 equal zones. The escape zone (ZC) contained the escape hole. We included an additional measure called the escape quadrant (ZQ), which contained the escape zone plus the two zones to the left and right (ZQ-2, ZQ-1, ZQ+1, ZQ+2) (Fig. 3b). Time spent in the escape zone and quadrant were calculated as well as latency to reach the escape hole, number of reference errors, hole deviations, spatial strategy used and distance traveled.
Fifteen min prior to the test, rats were infused with either saline (VEH) or PRO and returned to their home cage. Seven min prior to the test rats were given another infusion of either VEH or ISO. Infusion volume, rate, and procedure were the same as the previous infusion. Rats were then placed back in the home cage and tested 3 min later (each infusion took 4 min). This resulted in four groups: VEH-VEH, VEH-ISO, PRO-VEH and PRO-ISO. Following the acquisition, probe animals received 2 days of retraining (two trials per day, ITI = 2 h) to reduce any extinction learning that may have occurred during the acquisition probe (probe fixed in length; no escape box).
Curtain probeAnimals were re-trained following the acquisition probe and then given a curtain probe trial. The purpose of the curtain probe was to assess whether rats were using intra or extra-maze cues to locate the escape box. The procedure for this test was identical to the acquisition probe except that, animals did not receive any infusions and a brown plastic curtain was hung around the maze from the ceiling effectively blocking all visual access to the rest of the room. After this test animals received an additional 2 days of retraining (two trials per day; ITI = 2 h) to reduce any extinction learning that may have occurred during the curtain probe trial.
Reversal trainingSimilar to the acquisition probe, 1 h after the last retraining trial, stylets were unscrewed from each rat’s cannulae, and the infusion cannula was inserted and left in the guide cannula for 2 min on each side of the brain, stylets were then screwed back in and animals were returned to their home cage. The following day animals received their first reversal training trial.
Reversal training was similar to acquisition training except for the location of the escape box was moved 180 degrees. It lasted 5 days with one trial on the first day and 2 trials/day (ITI = 2 h) after that. Fifteen min prior to the first reversal training trial rats were given an infusion of either VEH or PRO. Seven min prior to RV1 rats were given another infusion of either VEH or ISO. Rats were placed back in the home cage and 3 min later tested on the reversal learning session. Groups were the same as the acquisition probe (VEH-VEH, VEH-ISO, PRO-VEH, PRO-ISO). Therefore, if a rat was in a specific group during the acquisition probe it remained in that group for RV1. One hour later, rats in the VEH-VEH group were split in half and were either returned to their home cage (VEH-VEH) or given an infusion of ISO (to form the new group VEH-VEH-ISO) and then returned to their home cage. This was to assess whether any effect of ISO was due to an enhancement of consolidation. The remainder of the reversal training trials occurred in the absence of any infusions.
Reversal probeFollowing reversal training, a reversal probe test was given to assess memory for the new escape hole location. The procedure was the same as the curtain probe but without a curtain. The same measures were recorded.
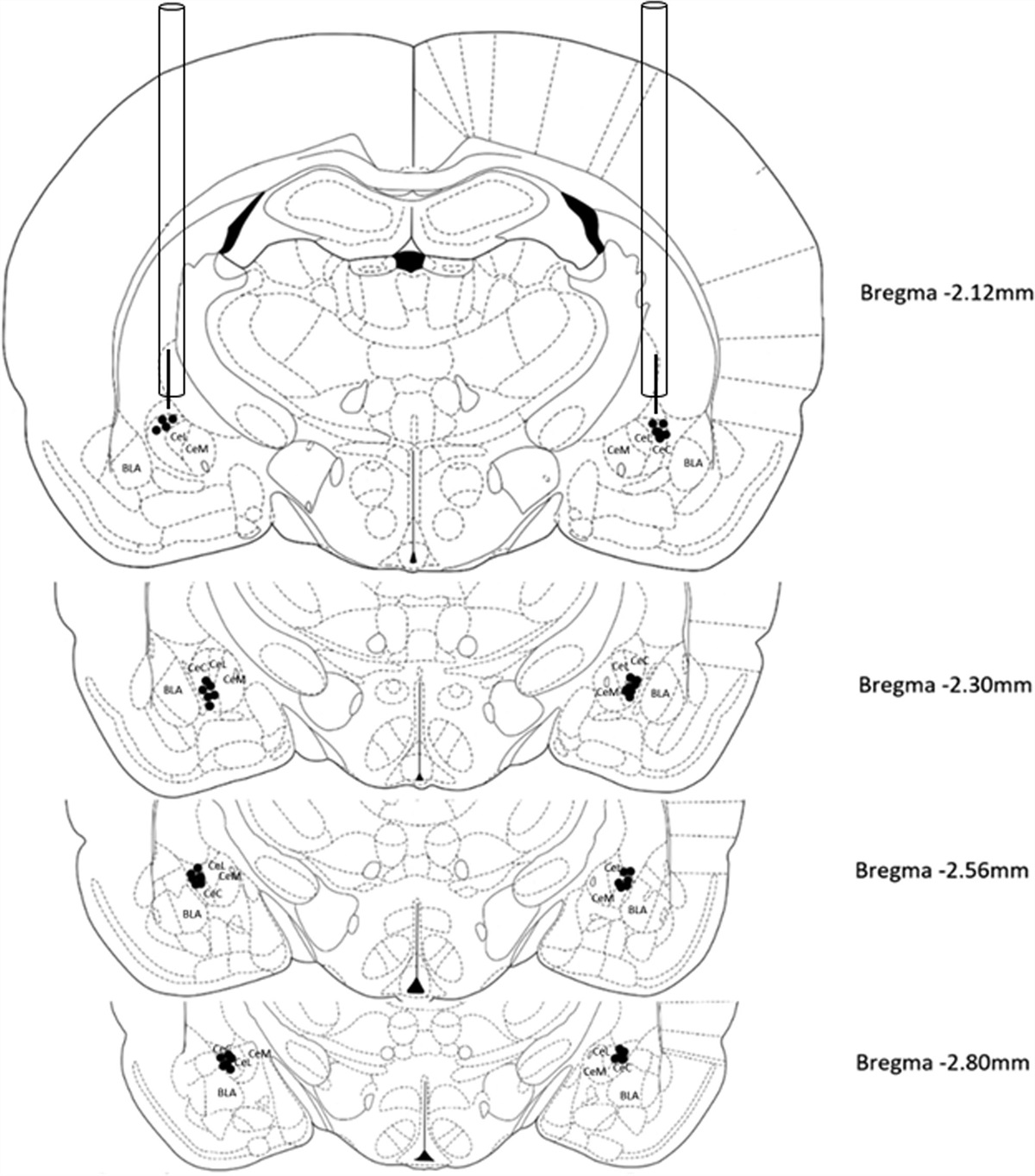
HistologyCannula placements were confirmed histologically at the end of the experiments. Following termination of behavioral experiments rats were transcardially perfused with cold 0.1M phosphate buffer solution (PBS) and subsequently cold 4% paraformaldehyde (PFA) in 0.1M PBS. Brains were left to post-fix for 1 h and then extracted and placed in PFA overnight. The following day they were transferred to a 30% sucrose/0.1M PBS cryoprotectant solution until saturated. They were then frozen and sectioned using a cryostat to produce 50 μm coronal sections. Every third slice was mounted onto gel-coated slides and Nissl-stained with Cresyl Violet. Slides were then cover slipped and cannula placements were verified under a microscope (Figure S1, Supplemental digital content 2, http://links.lww.com/BPHARM/A70). It should be noted that we did not verify the spread of the injections and therefore there is a small possibility that the drugs administered could have extended to adjacent brain regions such as the CA1-3.
Data collection and analysisStatistical analyses were conducted using SPSS (IBM, version 26) and SigmaPlotTM 11.0 (Systat Software, San Jose, California, USA). For the DNMP task, the dependent measures were latency to obtain reward, number of errors and the percentage of trials where a correct choice was made. Latencies were collected using a timer, and the experimenter recorded the number of errors, which was later used to calculate the percentage of correct trials. Pilot data were analyzed using one-way analysis of variance (ANOVA) to compare arm separations 1 through 6. Habituation and pretraining data were analyzed using two-way (GROUP x DAY) repeated measures ANOVAs. Acquisition data were analyzed using two-way (ARM-SEPARATION x DAY) repeated measures ANOVAs. Test data were analyzed using two-way repeated measures (GROUP x TRIAL) ANOVAs separately for each arm-separation. Washout and curtain probe data were analyzed using three-way repeated measures (GROUP x ARM-SEPARATION x DAY) ANOVAs. Pairwise comparisons were made when necessary, using Tukey’s HSD test.
In quantifying the EPM data, we measured distance traveled, mean speed, time spent immobile, line crossings, time spent in each zone of the maze and the number of entries into the zones. The locomotor measures (distance, speed, line crossings and immobility) were analyzed as one-way ANOVAs, and the time spent in each zone, as well as the number of entries, were analyzed using two-way (GROUP x ZONE) repeated measures ANOVAs. Pairwise comparisons were made when necessary, using Holm–Sidak tests.
For the Barnes maze data, we measured path efficiency, total distance traveled, latency to reach the escape hole, the number of hole deviations, reference errors and characterization of the search path used to find the escape hole. Using a Kruskal–Wallis test these data were compared across days during acquisition training. Difference scores in these measures were calculated between the acquisition probe and the last day of acquisition training. For each measure, one-way ANOVAs were then conducted to measure group differences. During each of the probe sessions, the maze was divided into 20 equal zones and the time spent in each zone was recorded. Group differences in the time spent in the escape zone, and the escape quadrant (which included the escape zone as well as the two zones to the left and right of the escape zone) were compared using a one-way ANOVA. To demonstrate whether differences in performance existed between the acquisition probe and the curtain probe a two-way (GROUP X SESSION) repeated measures ANOVA was conducted. On the first day of reversal training group comparisons in latency to reach the escape hole, hole deviations and reference errors were calculated using a one-way ANOVA. During the reversal probe, behavior across groups was analyzed with a one-way ANOVA. Pairwise comparisons were made when necessary, using Tukey’s HSD test. In all cases, P < 0.05 was accepted as significant. Error bars in graphs represent ± SEM, *P < 0.05.
Results Delayed nonmatch to position pilot experiment: how arm separations were determinedDuring the pilot experiment, rats were habituated to the maze and then taught to obtain a reward in one arm of the maze in under 2 min (sample trials). Once this behavior was acquired, they were given acquisition trials where they were required to remember the location of the previous arm that they had received that reward in (sample trial) and upon the presentation of two open arms, choose the arm they had not entered yet (choice trial) to successfully complete the task and receive an additional reward. Pilot animals were tested on choice trials where the arm separation ranged from 1–6. Across time, animals learned to obtain the reward more quickly (data not shown). On the last 3 days of training, there were no significant differences in the latency to obtain the reward across all arm separations (F5,66 = 1.65, NS) (Figure S2A, Supplemental digital content 2, http://links.lww.com/BPHARM/A70). However, there were differences in the number of errors made (F5,66 = 3.50, P < 0.01) and the percentage of correct trials (F5,66 = 3.18, P < 0.025) (Figures S2B,C, Supplemental digital content 2, http://links.lww.com/BPHARM/A70). Rats made more errors when the arm-separation was two arms compared to when the separation was five or six arms (2 vs. 5: P < 0.01; 2 vs. 6: < 0.05) (Figure S2B, Supplemental digital content 2, http://links.lww.com/BPHARM/A70). When the arm-separation was two arms, rats also had the lowest percentage of correct trials, which differed significantly from the 5-arm separation group (P < 0.025) (Figure S2C, Supplemental digital content 2, http://links.lww.com/BPHARM/A70) suggesting that the task was easiest when the separation was five arms and most-difficult when the separation was two arms. Given that Clelland et al. (2009) reported chance level performance when a one arm-separation (45 degrees) was used, and the fact that clockwise and counter-clockwise permutations cannot be counterbalanced for a 6-arm separation (180 degrees) in a 12-arm radial maze we decided to use the 2-arm separation for the difficult, DG-dependent, high similarity condition (S2, 60 degrees) and the 5-arm separation for the easier, DG-independent, low similarity condition (S5, 150 degrees). Moreover, these arm-separations were comparable to those used in Clelland et al. (2009) in terms of angular distance.
The dentate gyrus orthogonalizes contextual representations when they are highly similarIn the current study, we sought to assess the role of BAR activity during a spatial memory task that relied on the DG. It has been determined (Clelland et al., 2009), that spatial discrimination in an eight-arm radial maze is dependent on the DG when stimuli are presented with little separation but not when presented more widely apart. We adapted this task for the twelve-arm radial maze. Equating angular distance to obtain comparable behavioral results, we concluded optimal arm separations were two- and five arms apart since we saw the greatest difference in latency (Figure S2A, Supplemental digital content 2, http://links.lww.com/BPHARM/A70), number of errors (Figure S2B, Supplemental digital content 2, http://links.lww.com/BPHARM/A70), and percent of correctly conducted trials (Figure S2C, Supplemental digital content 2, http://links.lww.com/BPHARM/A70), demonstrating the S2 condition was more difficult and DG-dependent than the S5 condition. We then assessed the role of BAR activation and inactivation in the DG with ISO and PRO respectively, across both S2 and S5 conditions. During habituation, rewards collected increased across days (F3,114 = 9.42, P < 0.005) (Figure S2D, Supplemental digital content 2, http://links.lww.com/BPHARM/A70) with significantly more rewards collected on days 3 (P < 0.01) and 4 (P < 0.005) compared to day 1. During pretraining, only sample trials were presented. Animals initially took approximately 200 s to obtain the reward but by the 10th day, they could do this in under 30 s (F9,279 = 17.45, P < 0.005) with significantly shorter latencies emerging by pre-training day 3 (P < 0.005) (Figure S2E, Supplemental digital content 2, http://links.lww.com/BPHARM/A70). No group differences were observed.
During DNMP acquisition training, animals received both sample and choice trials. By the 6th day, 62.5% of the rats reached criterion; the other 37.5% reached this criterion by day 7. The analysis included the first 3 days and the last 3 days of training data; therefore, the data is inclusive for rats that took 6 days to reach criterion, and for rats that took 7 days, the set excludes the data from day 4. Animals completed the trials more quickly across time (F5,195 = 9.04, P < 0.005) and animals in the S2 condition took longer to complete the trials compared to the S5 condition (F1,195 = 19.38, P < 0.005) (Figure S2F, Supplemental digital content 2, http://links.lww.com/BPHARM/A70). At the start of acquisition training, all rats were performing at an error rate of approximately 50% (Figure S2G, Supplemental digital content 2, http://links.lww.com/BPHARM/A70). As they learned the task, a difference in performance emerged across groups (significant interaction: F5,195 = 4.37, P < 0.005) by acquisition day 5 (P < 0.025) with rats improving in the S5 condition to 94% correct trials on day 6 (P < 0.005) whereas this remained low in the S2 condition (Figure S2G, Supplemental digital content 2, http://links.lww.com/BPHARM/A70). A similar pattern emerged for the number of errors made in the choice trials (significant interaction: F5,159 = 2.49, P < 0.05) starting on day 5 (P < 0.01) and extending to day 6 (P < 0.005) (Figure S2H, Supplemental digital content 2, http://links.lww.com/BPHARM/A70).
Biasing memory towards encoding during retrieval can be maladaptive: pre-choice infusions of isoproterenol impaired choice accuracy and increased latency to obtain a reward in a working memory taskFollowing training, animals were tested on 4 conditions across four different testing days (Figure 1A, Table S6, Supplemental digital content 1, http://links.lww.com/BPHARM/A69). They were assigned to either the ISO or PRO drug group which remained consistent across all test days. Differing on each test day was the timing of the infusion: pre-sample or pre-choice (Fig. 1b), and the arm separation: 2 arms (S2) or 5 arms (S5) (Fig. 1c). Animals were given a habituation trial (HAB), a baseline trial (BASE; vehicle/saline), and a test trial (TEST, drug). On the basis of our hypothesis that NE biases the memory system towards encoding, we expected ISO given pre-sample would improve memory while PRO would have the opposite effect, in other words, biasing the system toward encoding when it is adaptive when learning is occurring. While we found no effect on latency or the number of errors during testing (Fig. 1d,f), we did find that infusions of PRO resulted in fewer trials where animals made the correct choice (F1,38 = 4.90, P < 0.05) potentially due to blockade of NE-driven encoding necessary to complete this task successfully (Fig. 1e).
Our hypothesis also led us to assume that ISO administered after the sample, and before the choice, when the task required recalling a previously formed contextual map rather than recruiting a new map, would result in impaired memory, in other words, biasing the system when it is maladaptive. As expected, animals in the S2 condition took longer than the S5 condition to obtain the reward (significant interaction: F1,38 = 8.42, P < 0.01). More specifically, animals in the ISO group demonstrated longer latencies on the test trial compared to baseline (P < 0.005) and compared to PRO animals (P < 0.005) in the S2 condition. In the S5 condition, ISO animals did not exhibit longer latencies during the test (Fig. 1g). One explanation for these results may be that ISO promotes attentional shifts where animals spend more time exploring the maze rather than focused on the task. While animals did make more errors (and fewer correct trials) when given ISO in both S2 and S5 trials (Fig. 1h,i), these effects were not statistically significant. However, this effect was more pronounced in the S2 condition demonstrating the DG is sensitive to this disruption.
Visuospatial learning in the delayed nonmatch to position task depends on contextual elements within the environmentSeparati
留言 (0)