
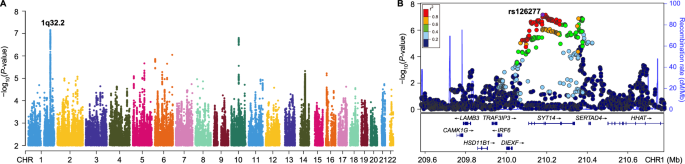
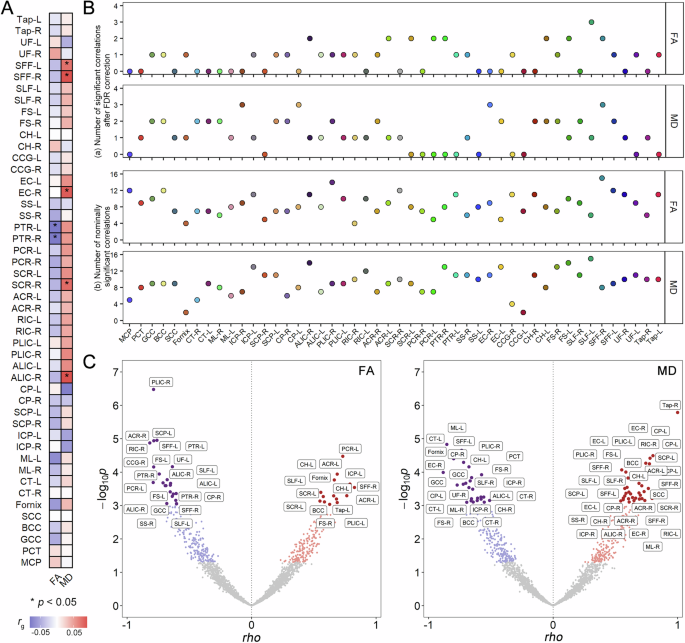
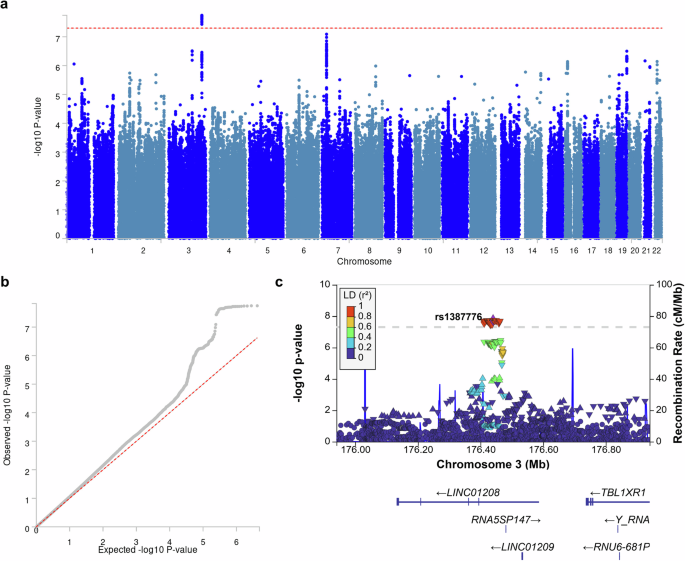
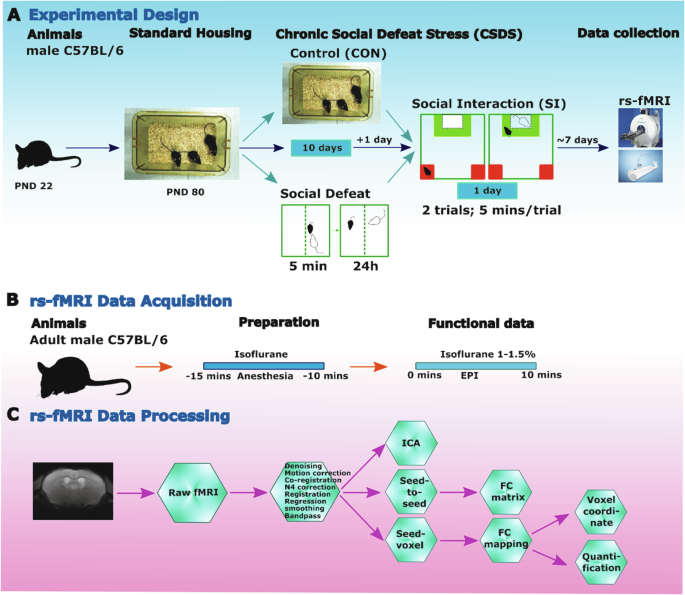
記住我


Increases in psychomotor activity and development of sensitization are behavioral phenomena caused by drugs of abuse and are mediated, at least in part, by alterations in dopaminergic neurotransmission [27, 28]. In naïve mice (N = 64, 30 male, 34 female), a single esketamine administration (10 mg/kg, i.p.) [4] produced a rapid and short-lasting increase in locomotion that was later followed by a decrease in activity below the levels of saline-treated mice (Fig. 1A–C, E). The statistical analysis of the distance traveled using a three-way ANOVA with repeated measures by subject with time bin, esketamine treatment and naloxone treatment as main factors showed significant effects of time bin (F(9.871, 641.6) = 34.81, p < 0.0001) and a triple interaction between time bin x esketamine x naloxone (F(29, 1885) = 2.241, p = 0.0002). At the same time, this effect was accompanied by a significant decrease in time spent in the center of the arena (Fig. 1A, B, F), a response indicative of increased anxiety-like behavior [29]. Three-way ANOVA with session, esketamine and naloxone as factors showed significant effects of esketamine (F(1, 60) = 4.716, p = 0.033), but no interaction between esketamine and naloxone. In summary, while the initial locomotor stimulating effect was partially blocked by the mu-opioid receptor (MOR) antagonist naloxone (3 mg/kg, s.c.), both the late locomotor depression and the center time were not (Fig. 1C, F).
Fig. 1: Some, but not all, psychomotor effects of esketamine require opioid receptor engagement.
Representative trajectory of a representative mouse placed in an arena for 30 min after saline A or acute esketamine administration B, where the color of the line depicts the speed of the animal. C and D Time course of the elapsed distance before and after saline (grey), naloxone (blue), esketamine (orange) or naloxone and esketamine (green) administration, line and shade represent mean ± SEM of all animals. E Accumulated distance during the 10 min following drug (or saline) administration (each dot represents one animal, bars represent mean, summary statistics: post-hoc Tukey’s multiple comparisons test: *p < 0.05, ** p < 0.01). F Percentage of time the animals spent in the center of the arena (each dot represents one animal, summary statistics: post-hoc Tukey’s multiple comparisons test: *p < 0.05, ** p < 0.01) G Representative heatmap of the preferred locations of mouse placed in the Elevated Plus Maze for 5 min after saline or acute esketamine administration. H, I Time spent in the open arms, and total distance traveled by the animals (each dot represents one animal, statistical significance calculated by t-tests: ***p < 0.001 ****p < 0.0001.
We continued to inject the mice with esketamine (10 mg/kg, i.p., daily) for four more consecutive days and on the fifth day we re-evaluated the acute effects of the drug. In agreement with our previous results [4], we found that esketamine increased locomotor activation in male mice on Day 5 compared to Day 1, a response indicative of sensitization (Fig. 1E, S1) (main effect of session: F(1, 26) = 8.510, p = 0.0072), however female mice did not show increased locomotion after repeated esketamine administration (Fig. S1) (session: F(1, 28) = 0.8347, p = 0.37). Interestingly, on male mice while the statistical analysis of the time course (Fig. 1D, S1) showed a triple interaction between time bin, esketamine and naloxone on Day 1 (F(29, 1885) = 2.241, p = 0.0002), there was no interaction between these factors on Day 5 (F(29, 1766) = 0.5925, p = 0.95). On the other hand, and in contrast to the first esketamine administration, esketamine did not change the time spent on center after 5 days of repeated administration (Fig. 1F) and the statistical analysis showed an interaction between esketamine and experimental session (F(1, 55) = 22.32, p < 0.0001). Overall, these data suggest that single vs. repeated esketamine administration produces multifaceted effects on both locomotor activity and anxiety-like behavior, with the locomotor stimulating effects being dependent on opioid receptors.
To further confirm the putative anxiogenic-like effects of acute esketamine administration we performed the elevated plus maze (EPM) test [30] in naïve mice (N = 78, 42 male, 36 female). Mice received saline or esketamine (10 mg/kg, i.p.) and 15 min later they were placed in the center of an EPM and their trajectory was evaluated for 5 min (Fig. 1G). Male mice injected with esketamine spent significantly less time exploring the open arms of the maze (Fig. 1H) and this effect was not affected by pretreatment with naloxone. However, female mice -which on saline conditions spent less time exploring the open arms than male mice- were unaffected by esketamine administration. Three-way ANOVA with sex, esketamine, and naloxone administration as factors revealed significant main effects of sex (F(1, 69) = 4.260, p = 0.0428) and esketamine (F(1, 69) = 6.240, p = 0.0149) and an interaction between sex and esketamine (F(1, 69) = 18.50, p < 0.0001). However, consistently with the open field data, although both sexes showed similar overall activity upon vehicle administration, only male mice were less active when administered with esketamine and this effect was blocked by naloxone (Fig. 1I). Three-way ANOVA: main effect of esketamine (F(1, 72) = 12.97, p = 0.0006), interaction naloxone x esketamine (F(1, 72) = 2.947, p = 0.0904), sex x esketamine (F(1, 72) = 5.691, p = 0.0197). To try to correct for this putative biasing factor, we calculated the relative distance traveled in the open arms (Fig. S1). These data showed no significant effects on esketamine-treated mice (Fig. S1, F(1, 73) = 2.645, p = 0.1082) and hence indicated that the avoidance of the open arms was not due to an impairment of their locomotor performance or sedation, but active avoidance of the open arms, confirming that a subanesthetic dose of esketamine increases anxiety-like behavior in male mice.
Esketamine alters phasic dopamine frequency and re-uptake kinetics in miceThe behavioral data presented above indicated that esketamine produces behavioral effects that are typically modulated by mesolimbic dopaminergic neurotransmission. Therefore, to explore the potential involvement of dopamine in the pharmacological effects of esketamine, we used the genetically encoded dopamine biosensor dLight1.3b [31] to monitor extracellular dopamine concentrations in the NAc of mice (Fig. 2A). Three to four weeks after intracranial injection of a viral vector AAV-1/2-hSyn1-Dlight1.3b in the NAc of mice (N = 35, 18 male, 17 female), we inserted a fiber optic into the injection site and fixed it to the skull (Fig. 2A, B). After 4–5 days of recovery, we recorded the dopamine-elicited changes in fluorescence while the animals were in their home cage after an acute administration of saline, naloxone (3 mg/kg, s.c.), esketamine (10 or 30 mg/kg, i.p.) or naloxone plus esketamine (Fig. 2C–G, Fig. S2). Esketamine did not produce any observable change in dopamine tone (Fig. 2C, Fig S2), however the 30 mg/kg dose of esketamine decreased the frequency of dopamine transients (Fig. 2E; mixed-effects model with repeated measures (REML) with esketamine, naloxone and session as factors: main effect of esketamine (F(2, 36) = 11.09, p = 0.0002) and it was unaffected by naloxone (REML interaction esketamine x naloxone: F(2, 36) = 0.07,147, p = 0.93)). Esketamine administration also produced a dose-dependent change in the kinetics of the spontaneous dopamine transients compared to vehicle-treated mice and this effect was blunted upon naloxone pre-administration. Specifically, while the effects on dopamine peak amplitude were not altered (Fig. 2F, Fig. S2), the duration of the dopamine transients increased significantly from 0.96 ± 0.09 s in baseline conditions to 1.17 ± 0.11 and 1.30 ± 0.13 after 10 and 30 mg/kg esketamine, respectively (Fig. 2G, Fig. S2; REML esketamine: F(2, 34) = 25.41, p < 0.0001; esketamine x naloxone: F(2, 34) = 5.035, p = 0.0122). Treating mice with vehicle or naloxone at the same time intervals as esketamine did not alter the photometry signal during the course of the experiment, indicating that the observed effects were due to esketamine administration and not time spent in the testing arena (Fig. S2). These results indicate that the so-called increase in dopamine neurotransmission caused by esketamine is not due to an enhancement of dopamine release but to longer duration of dopamine presence in the postsynaptic area.
Fig. 2: Esketamine administration causes mixed effects on spontaneous dopaminergic transmission.
A Schematic representation of dopamine sensor expression in the NAc. B Representative immunohistochemical image of dLight1.3b expression in the NAc. Scale bar: 1 mm. C Average trace detected from the fluorescent signal of dLight1.3b in the NAc after saline or esketamine (10 mg/kg and 30 mg/kg) administration. D Representative traces of the corrected dLight1.3b in the NAc after saline or esketamine (10 mg/kg and 30 mg/kg) administration. E–J Peaks per minute, dopamine transient amplitude and dopamine transient duration on day 1 and day 5 of esketamine administration with and without pretreatment with naloxone (3 mg/kg) (each dot represents one animal). K Schematic representation of the viral strategy for expression dopamine and calcium biosensors in the NAc and VTA-originated terminals, respectively, and representative traces of Dlight1.3b, RCaMP1.07 and isosbestic control signals. L–N Peaks per minute, transient amplitude, and transient duration of calcium in the NAc (each dot represents one animal). In all panels, statistical significance, evaluated by Tukey’s post-hoc multiple comparisons test, * p < 0.05 and ** p < 0.01). O, P Correlation between extracellular dopamine and intracellular calcium from terminals from VTA neurons in the NAc. All correlations were statistically significant with p < 0.001 and esketamine treatment did not alter Pearson’s r coefficients on amplitude (p = 0.52) nor duration (p = 0.58).
We then repeated the same experiment after four days of daily esketamine injections (Fig. 2H–J). Before esketamine administration, dopamine transient frequency and their amplitude and duration did not differ from naïve mice (Fig. 2H–J). However, esketamine administration after four days had weaker effects on frequency and transient duration, but increased the dopamine transient amplitude, an effect that was not observed on day 1 (Fig. 2H–J). Statistical REML analysis showed a significant interaction between esketamine and session on transient duration F(2, 15) = 3.921, p = 0.042) and no significant effects of interaction between naloxone and session were observed in any of the three parameters.
We aimed to evaluate whether the esketamine-induced extracellular dopamine increases were due to pre- or postsynaptic actions of esketamine. In a subset of mice (N = 5 male, 1 female), we injected a viral vector encoding the red fluorescent calcium biosensor RCaMP1.07 (AAV-1/2-hSyn1-RCaMP1.07) into the VTA together with the dLight1.3b sensor in the NAc. In these mice, NAc dLight1.3b (from postsynaptic cell bodies and dendrites) and RCaMP1.07 fluorescence (from dopaminergic presynaptic terminals) were recorded simultaneously to establish a correlation between the two signals (Fig. 2K). We observed a high coherence between calcium and dopamine signals (Fig. 2K, O, P) and, in agreement with the dLight1.3b data, esketamine increased the duration of the spontaneous calcium transients in NAc dopaminergic terminals (mixed-effects model with repeated measures: F(1.733, 6.066) = 13.68, p = 0.0063) without affecting their amplitude (Fig. 2L–N).
Esketamine increase in electrically evoked dopamine is dependent on MOR engagementTo explore the pharmacological mechanisms underlying esketamine’s effects on dopamine neurotransmission we used local electrical stimulation to induce neurotransmitter release in anesthetized mice [32] (N = 32, 20 male, 12 female). This protocol allowed repeated and controlled release events and hence it allowed us to test drug actions over time after establishing a baseline for each mouse (Fig. 3A, B). In concordance with the awake photometry data (Fig. 2), esketamine increased electrically evoked dopamine peak duration (Fig. 3C, D; two-way ANOVA with esketamine dose and presence or absence of naloxone (see below), main effect of esketamine: F(2, 36) = 68.06, p < 0.0001) without altering peak amplitude (Fig. 3E; two-way ANOVA (esketamine’s main effect): F(2, 37) = 0.07,698, p = 0.9261)), and resulting in an overall increase in extracellular dopamine concentration as quantified as the area under the curve (AUC) of each peak (Fig. 3F; two-way ANOVA (esketamine’s main effect): F(2, 32) = 81.23, p < 0.0001).
Fig. 3: Multiple molecular mechanisms account for the dopaminergic effects of esketamine.
A Schematic representation of AAV injection site and fiber/electrode placement in the NAc. B Representative heat map of extracellular dopamine concentrations before and after each electrical stimulation event. C Representative dopamine traces before (black) or after esketamine administration (10 mg/kg, orange, 30 mg/kg purple). D–F Duration, amplitude, and AUC of extracellular dopamine transients. G Schematic representation of AAV-hSyn-GCaMP7s injection site and fiber/electrode placement. H Representative calcium (GCaMP7s) traces after esketamine administration (10 and 30 mg/kg). I–K Duration, amplitude, and AUC of the electrically-evoked intracellular calcium transients. L Representative traces of extracellular dopamine (RdLight1) after optogenetic stimulation of the ChR2-expressing terminals (red) or non-ChR2 expressing terminals (black) in the NAc. Cyan line represents the stimulation period (400 ms). M Schematic representation of AAV-RdLight1 injection in the NAc, Channelrhodopsin-2 (ChR2) injection in the VTA and fiber in the NAc that was used for both recording and stimulation. N Representative RdLight1 trace after several optogenetic (cyan) and electrical (black) stimulation events. O Averaged traces (3–6 per animal) of electrical or optogenetic stimulation events before or after esketamine (30 mg/kg) administration. In all panels, each dot represents the average of 3 to 5 trials per animal and the bars represent the mean of all dots; statistical significance, evaluated by Tukey’s post-hoc multiple comparisons test, * p < 0.05, ** p < 0.01, **** p < 0.0001).
We used the mu-opioid receptor (MOR) antagonist naloxone (3 mg/kg, s.c.) to evaluate whether the dopaminergic effects of esketamine were dependent on MOR. Naloxone partially blocked esketamine-induced increases in peak duration and AUC (Fig. 3D–F). Two-way ANOVAs with esketamine dose and presence or absence of naloxone showed a significant interaction between esketamine and naloxone for peak duration (F(2, 37) = 0.4700, p = 0.6287) and AUC (F(2, 32) = 13.10, p < 0.0001). However, the opioid agonist morphine (10 mg/kg, i.p.) did not produce any dopamine effects using this preparation (Fig. 3D–F), indicating that MOR engagement alone is not sufficient to cause an increase in electrically-evoked dopamine release. We also compared ketamine to the high affinity and selective NMDAR non-competitive antagonist phencyclidine (PCP), which has 10-times higher affinity for NMDAR and >1000-times selectivity over opioid receptors [4, 5, 11, 33]. An equipotent dose of PCP (3 mg/kg, i.p.) produced a similar effect to esketamine (Fig. 3D–F), however this effect was not blocked by naloxone. To test whether these effects were due to prolonged activation of the dopaminergic terminals, we used the green calcium sensor GCaMP7s injected in the VTA and we placed a fiber optic/electrode in the NAc (Fig. 3G, H). Upon esketamine administration, we did not observe any quantitative difference in the main intraterminal Ca2+ increase (Fig. 3I–K), however we observed that the calcium dynamics traces have two components, one main increase event (caused by the electrical stimulus) followed by a secondary minor increase (Fig. 3H). Unfortunately, the location of the secondary increase was inconsistent between animals and oftentimes merged with the main release event and could not be quantified.
Overall, these data indicate that the electrically-evoked increases in dopamine are not mediated by a single pharmacological mechanism but that several neurochemical systems are involved. The method used to electrically evoke dopamine release necessarily produces local release of other neurotransmitters, which might in turn locally affect dopaminergic cell activity [34, 35]. Therefore, we used an optogenetic approach to selectively activate NAc dopaminergic terminals. We injected Channelrhodopsin-2 (ChR2) in the VTA, and the red fluorescent dopamine biosensor RdLight1 in the NAc. Then, we lowered an optic fiber into the NAc that was used both to stimulate ChR2 (470 nm, 50 Hz during 400 ms) and simultaneously record red fluorescence from RdLight1 (Fig. 3L, M). A bipolar electrode was also attached to the optic fiber, allowing us to compare electrical vs optogenetic stimulation (Fig. 3N) with alternating pulses in the same mouse. Blue-light stimulation produced transient increases in RdLight1 signal only in animals transduced with ChR2, but not in control mice (Fig. 3L), confirming that the signal represented dopamine release rather than artefactual blue-light excitation of this biosensor as recently described [36]. While the amplitude of the peaks generated by either electrical or optical stimulation was comparable in scale factors, the duration of the ChR2-induced peaks was markedly reduced (Fig. 3N, O), suggesting that the release of other neurotransmitters caused by electrical stimulation sustains the duration of the dopamine release. Esketamine (30 mg/kg) did not alter the dynamics of the optogenetic dopamine transients (Fig. 3O). Taken together, these results suggest that esketamine modulation of dopamine neurotransmission does not occur by direct modulation of the NAc dopaminergic neurons but rather by affecting other neurotransmitters locally in the NAc.
Esketamine rapidly decreases glutamatergic transmission in the NAcTo test whether alterations in glutamate release might contribute to the dopaminergic effects of esketamine, we injected mice with a mix of the green fluorescent glutamate sensor iGluSnFR and the red dopamine sensor RdLight1 (Fig. 4A, Fig. S3) in the NAc. In one set of mice (N = 6, 4 male and 2 female), we performed electrical stimulation under anesthesia using the same conditions described previously (Fig. 4B). Acute esketamine administration (10 or 30 mg/kg, i.p.) did not produce a significant dose-dependent effect on glutamate release (Fig. 4C–E). We then performed fiber photometry on awake free behaving mice. Mice (N = 5, male) were allowed to explore their home cage for 20 min and, after that, 10 and 30 mg/kg of esketamine was injected at 30 min intervals. In contrast to the data using the green dopamine biosensor dLight1.3b (Fig. 2C), we observed a fast increase in dopaminergic tone and a decrease in glutamatergic tone after esketamine injection (Fig. 4F, G). In addition, the peak analysis found a reduction on glutamate transient frequency (RM one-way ANOVA, (F(1.753, 7.010) = 6.654, p = 0.026; Fig. 4H) but no effect on the duration of individual glutamate transients (RM one-way ANOVA: F(1.353, 5.411) = 0.4576, p = 0.58; Fig. 4I) nor their amplitude (RM one-way ANOVA:; Fig. 4J). Taken together, these results suggest that the increase in esketamine-induced increase in dopamine activity in the NAc is accompanied by a decrease in glutamatergic activity.
Fig. 4: Esketamine has opposite effects on dopaminergic and glutamatergic transmission in the NAc.
A Schematic representation of dopamine and glutamate sensors injected in the NAc. B Representative traces of electrically evoked extracellular dopamine (red) and glutamate (green) in the NAc. C–E Amplitude, duration and AUC of glutamate release upon electrical stimulation after administration of saline (black) or esketamine 10 mg/kg (orange) or 30 mg/kg (purple). F Average of extracellular glutamate (green) and dopamine (red) traces before and after esketamine administration (upper panel), or after saline administration (bottom panel). Each line is the average of 4–5 animals. G AUC of the tonic changes in dopamine (red) or glutamate (green) during 20 min after drug administration. Each dot represents one animal, and the bars represent mean ± SEM. H–J Quantification of the glutamate transients (frequency, amplitude, and duration). In all panels except (F) and (G), each dot represents the average of 3 to 5 trials per animal and the bars represent the mean of all dots; statistical significance, evaluated by Tukey’s post-hoc multiple comparisons test, * p < 0.05, ** p < 0.01, *** p < 0.001).
Reward-evoked dopamine release is impaired by acute esketamine administrationThe data presented above suggests that esketamine alters basal and stimulated dopaminergic activity. Dopamine is a key modulator of motivated behaviors [37]. Therefore, we examined the relationship between esketamine-induced increases in dopamine activity and motivated behavior. Mice (N = 24: 12 male, 12 female) were trained to nose poke on a port to obtain a delivery of a 20 mg sucrose pellet under increasing fixed-ratio (FR) schedules (1, 5 and 10). Nose poking on an inactive second port did not result in any pellet delivery. Once their responding on FR was stable (see Methods) they were moved to a progressive-ratio (PR) schedule, in which the number of pokes required to obtain a pellet exponentially increased after each pellet delivery within a session (Fig. S4). After 3 days on PR schedule to establish a baseline response, animals were administered with esketamine (10 mg/kg, i.p.; Fig. 5A, B) five minutes before the start of the session. One subset of mice was also pretreated with naloxone (3 mg/kg, i.p.) 10 min before esketamine administration. Female mice (Fig. A, C) reached a higher break point than male mice (Fig. 5B, D), but when the data were normalized to the rewards obtained under baseline conditions, no statistical differences by sex were observed and, therefore sex was not considered a statistical variable. Esketamine treatment reduced the total number of rewards obtained (Fig. 5E) and increased the time it required to achieve them (Fig. 5F), however naloxone did not block these effects. Esketamine effects were most pronounced during the first 20 min of the session, a time point corresponding to esketamine-induced hyperlocomotion (Fig. 1) and tonic changes in glutamate and dopamine (Fig. 4). To test whether their ability to respond correctly or retrieve pellets was impaired, we measured the ratio of correct responses and the time elapsed between pellet delivery and retrieval. Esketamine reduced the ratio of correct responses (Fig. 5G; REML showed a positive interaction between time bin and treatment: F(10,84) = 3.73, p = 0.0004), and also increased the pellet retrieval time (Fig. 5H; Brown-Forsythe ANOVA 12.21 F(4.000, 417.7) = 12.21, p < 0.0001).
Fig. 5: Esketamine administration impairs reward-evoked dopamine release.
A–D Cumulative sucrose rewards obtained on a PR schedule after saline (black), esketamine (orange) or esketamine and naloxone (green) administration on days 0 (black) 1 and 5 of the procedure 5 days. E Total rewards obtained in a 1-h session for each depicted condition (each dot represents one animal, bars represent mean ± SEM of all animals). F Time in minutes the animals need to obtain half of the total amount of pellets (each dot represents one animal; floating bars represent 95% confidence intervals). G Time course of the percent of correct pokes in 10-min time bins. Each dot represents mean ± SEM of 12 animals per group. H Delay (s) between pellet delivery and pellet retrieval by the animal. Each dot represents one animal. I Representative trace of dLight1.3b and isosbestic control signals in the NAc during sucrose self-administration on FR5. Triangles represent animal actions: rewarded active poke (green), inactive poke (red) or pellet retrieval (black). J Heatmap representing extracellular dopamine variation on each pellet delivery event (aligned to the onset of the cue -black triangle-). Each row is one individual trial from 9 different animals (5–10 trials per animal). K Average of the corrected dLight1.3b signal aligned to the cue (top) or pellet retrieval (bottom) in control (black) or esketamine (10 mg/kg)- treated animals. Each trace is the average of 9 animals. L–N Amplitude, duration and AUC of the dLight1.3b signal increase during cue presentation, each dot represents the average of all trials (5–10) per animal (9) on saline of esketamine sessions. In all panels statistical significance, evaluated by the corresponding test (see main text): * p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001).
To evaluate dopamine dynamics during sucrose self-administration, we injected mice (N = 9 male) with the dopamine biosensor dLight1.3b in the NAc and performed fiber photometry while they performed sucrose self-administration on an FR5 schedule (Fig. 5I). The conditioned stimulus that signaled reward delivery elicited a transient dopamine increase (Fig. 5J, K) that lasted 1.1 ± 0.5 s (Fig. 5K, L). In the following session the mice were pretreated with esketamine (10 mg/kg, i.p.) five minutes before the start of the session. The mice that were administered esketamine also showed a transient increase in extracellular dopamine upon achievement of a reward (1.2 ± 0.5 s, paired t(8) = 0.81, p = 0.44, vs Saline condition). However, the amplitude of the dopamine transient was reduced by ~50% (ratio paired t(8) = 4.54, p = 0.0019, Fig. 5M). Consequently, the AUC of this dopamine transient in the animals treated with esketamine was also reduced by ~65% (ratio paired t(8) = 4.88, p = 0.0012, Fig. 5N). Overall, this data suggests that despite the higher dopaminergic activity elicited by esketamine in other experimental procedures, esketamine impairs reward-evoked dopaminergic activity.
留言 (0)