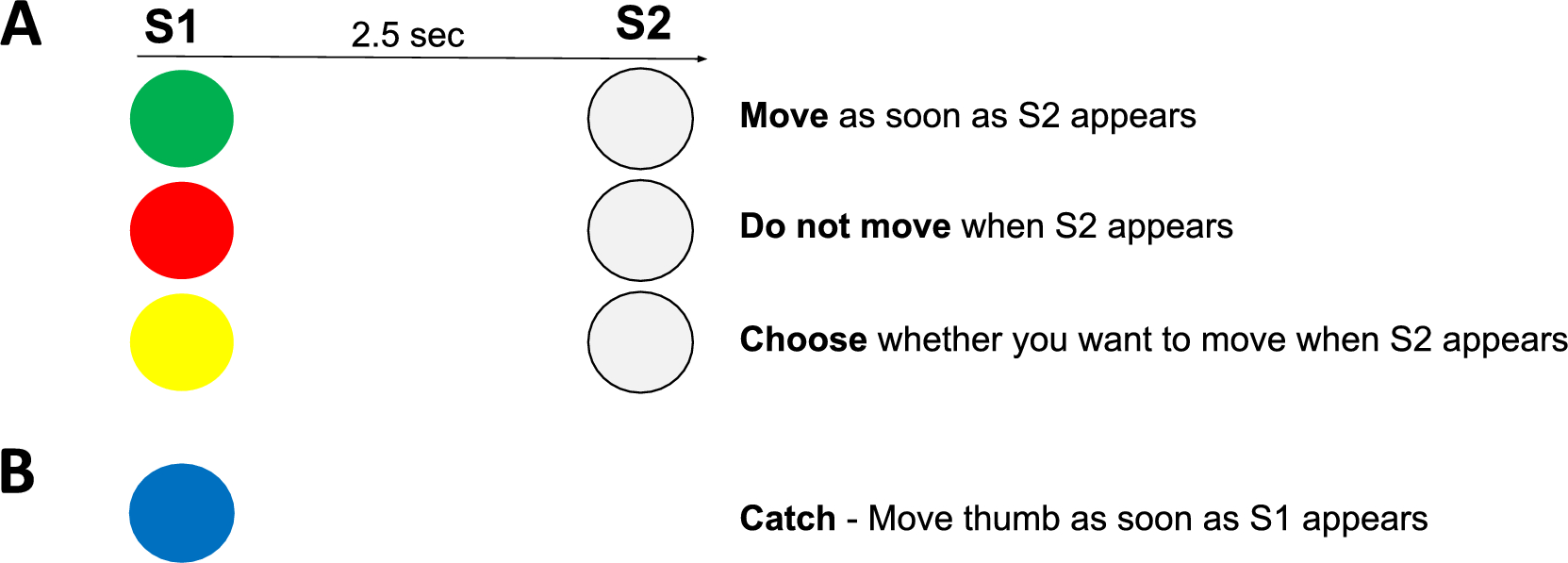
We used the CNV to look for signs of the decision making process for a free-choice decision to move (go) or not move (no-go) compared to that of commanded action or inaction. We found that free-choice and command CNVs were similar with the free-choice records lagging the command CNVs by short amounts of time. Thus, we see that when there is about 2500 ms to make a decision it takes about 1.5–2 s to do so. As an unexpected finding, we found that the CNV demarcation of a decision was “late” compared to much earlier apparent decisions in other parts of the brain. Thus, it appears that the CNV decision is actually more likely implementation of a movement decision that is made earlier.
Here we show that the CNV for command and choice tasks share similar responses in terms of amplitude at the time immediately before action, but differences in activity during the period leading up to this moment. The exploratory observations identify evidence that brain activity underlying beta modulation shows early brain activity that appears to play a significant role in the decision-making process well before the decision declares itself in the CNV. Our findings support the idea that initiating and canceling an action are closely related.
Development of CNV
We found our a priori hypothesis to be true that there was a significant difference in the CNV amplitude in the 50 ms prior to S2 between the choice go and choice no-go conditions as well as the command conditions. In addition, we found that choice go and command go conditions had similar amplitude, as well as the choice no-go and command no-go conditions.
The post hoc inspection of the CNV waveforms over time seem to reveal patterns regarding the decision-making process. The relatively long interval between S1 and S2 allowed us to visualize the brain’s response to a commanded condition versus one in which a decision could be made about whether to move or to not move (Figs. 2, 3). Starting with S1, we first see the response to the visual cue in the first 1000 ms of the CNV waveform. At about 1000 ms we see that the command go conditions and the command no-go conditions begin to diverge, but this does not reach significance until 1600 ms (900 ms before movement). After divergence, the command no-go condition remains close to 0 µV and the command go condition moves toward a voltage of about − 5 µV.
During the choice tasks, we find that the CNV waveforms remain together and with a voltage in between the command go and no-go conditions, suggesting that a decision has not yet been made. The divergence between these conditions is in the same direction as the command conditions but happens much later, at approximately 2150 ms after S1 (350 ms before movement). Our findings of the CNV and alpha and beta ERD in the command conditions are similar to previous findings (Filipovic et al. 2001; Filevich et al. 2012), but there have not been previous studies with choice conditions.
Regarding the CNV, an apparent conclusion is that the decision for command conditions is made at 1600 ms after S1, and the decision in the choice conditions is not made until 2150 ms after S1. The exploratory beta topoplots, however, show the decision to be much earlier, and moreover, show how the brain goes from decision to action (Fig. 4). Hence, the CNV more likely indicates movement preparation than the decision process. We acknowledge that some participants might make their choice at S2 rather than at S1, as suggested by studies from Kuhn and others (Kuhn et al. 2009). This could contribute to the later CNV divergence observed between go and no-go trials for the free choice condition. To understand our results, it is necessary to review prior studies of go/no-go experiments with and without choice.
Prior literature
There have been many prior studies using different variations of a go/no-go paradigm, mostly with fMRI. As the brain areas for movement are well known, the main interest in these studies is what happens in no-go situation. Results from all studies are consistent in showing areas that are more active with no-go than go indicating that inhibition is an active process. The specific areas vary to some extent among the studies, likely due to variations in the paradigms, but there is some consistency. The lateral part of the prefrontal cortex, either right (Dunovan et al. 2015; Aron et al. 2005) or left (Criaud et al. 2013; Brass et al. 2007; Kuhn et al. 2009) or bilateral (Watanabe et al. 2002) is most commonly identified. This region is described as inferior frontal gyrus (IFG), mid-frontal gyrus (MFG), left dorsal frontomedian cortex (dFMC), or dorsolateral prefrontal cortex (DLPFC). The pre-supplementary motor area (preSMA) is frequently noted (Dunovan et al. 2015; Simmonds et al. 2008), and sometimes premotor cortex (PMC) (Watanabe et al. 2002). One study noted directed connectivity from the left dorsal MFG to the preSMA (Kuhn et al. 2009). The inferior parietal lobule (IPL) is also frequently noted, left (Criaud et al. 2013; Watanabe et al. 2002; Brass et al. 2007) more often than right (Kuhn et al. 2009). These studies have identified important structures, but do not reveal their time course. Other than the two studies noted above that looked at the CNV with command instructions, there are no EEG studies. Takeyama et al. (Takeyama et al. 2022) studied no-go event-related potentials (ERPs) with intracranial EEG (ECoG) and found activity in the left posterior MFG and in the preSMA, SMA, and PMC regions. They then stimulated these areas and found inhibitory effects.
Recent studies have further explored the impact of varying Go/No-Go ratios on inhibition-related brain activity. For example, an event-related potential (ERP) study found that different ratios of Go and No-Go trials significantly influence behavioral performance and brain activity related to response inhibition. As the proportion of Go trials decreased, behavioral performance in Go trials improved, and error rates in No-Go trials decreased, highlighting the modulatory effect of trial ratios on inhibition-related neural processes (Zhang et al. 2024).
Furthermore, recent research has highlighted the differences between Go/No-Go and Stop-Signal tasks in terms of their neural and cognitive mechanisms. While both tasks measure inhibitory control, Go/No-Go tasks are more reflective of automatic inhibition, whereas Stop-Signal tasks are indicative of controlled inhibition. Neuroimaging studies have shown that these tasks engage overlapping but distinct neural circuits (Sebastian et al. 2013). Additionally, cognitive studies have shown that external factors, such as negative stimuli, can differentially modulate performance in these tasks, further supporting the idea that they rely on different inhibitory processes (Littman and Takács 2017).
There are fewer fMRI studies dealing with free choice in a go/no-go paradigm. Si et al. (2021) reported a meta-analysis that revealed bilateral anterior cingulate and preSMA, bilateral prefrontal cortex (PFC), bilateral IPL, and right PMC. Kuhn et al. (2009) found specific activation in anterior cingulate. EEG was reported in such a free choice experiment also (Leocani et al. 2001), but data were not analyzed to identify the pattern of relevant regions. In many other types of free choice paradigms, the mesial motor areas are typically involved. The conclusion is that there are many studies, but most deal with neuroimaging, and the time course of events is difficult to discern with that modality.
Limitations
A limitation of our study is the small sample size, which may reduce statistical power and increase the risk of Type II errors. This could lead to an underestimation of the true effects or failure to detect smaller but meaningful differences. However, this sample size is consistent with other similar studies in the field (e.g., do Nascimento et al. 2006; Brass and Haggard 2007; Shakeel et al. 2015). Future studies with larger cohorts are needed to confirm and extend these findings.
Due to the exploratory nature of our analysis and the limited number of trials, we were unable to perform a statistical analysis of the ERD topoplots. The complexity of the variables—frequency, time, channel, and condition—further complicated the analysis. As a result, our findings regarding the ERD are qualitative in nature. Future studies with a more focused scope and increased trial numbers may allow for a more detailed statistical exploration of these phenomena.
This study was initially designed to focus on the CNV, with the analysis of full brain activity included as an exploratory component. We had no prior hypothesis about the specific value of the beta activity, which unexpectedly emerged as particularly informative. Further experiments of this type, with better quantification and variations in movement tasks, would be valuable. Collecting sufficient data for each participant would also enable the examination of individual differences, which could provide additional insights.
Additionally, we were unable to perform source analysis and instead relied on prior literature to infer the brain areas most active in tasks of this type. Conducting the same experiments with fMRI in the same participants would help to refine our understanding of the anatomical locations involved in these processes.






留言 (0)