Brain signal variability is thought to be crucial for optimal cognitive and behavioral function [27] and is theorized to support the dynamic interplay between integrated and segregated brain states, which allows different brain regions to synchronize or operate independently depending on the specific task requirements [27]. Previous studies have focused on BOLD fMRI variability within the context of aging [6, 7, 28] and neurodevelopmental disorders [5, 13]. The current study investigated the age-related relationship between resting-state BOLD fMRI variability and repetitive behaviors in ASD in a large publicly available sample for the first time. The results show that rMSSD can differentiate between ASD and TD individuals using approaches that identify group differences with and without consideration of age. The results also show that restrictive and repetitive symptom severity in ASD as measured by the ADI-RRB scale is associated with rMSSD in both a linear and quadratic manner with age. Finally, controlling for demographics such as IQ, handedness, and gender in a subset of individuals did not affect the relationship between rMSSD and ADI-RRB scores. It also increased the spatial extent of categorical group differences, age x group, quadratic age x group effects, and the quadratic age x RRB interaction. Taken together, these results show how rMSSD may be used to identify differences in BSV between individuals with ASD and TD and how rMSSD may be used to identify associations with restrictive and repetitive behaviors in individuals with ASD.
Group differences in brain signal variability
This pattern of BSV in TD individuals generally aligns with earlier investigations reporting a general linear decrease in BSV across the cortex in TD individuals across the lifespan [4]. This general decrease is thought to be associated with maturational processes that occur with the degradation of processing speed and cognitive abilities across old age [6, 9, 29]. However, caution should also be used when comparing the results of the current study against previous research, as the age ranges differ considerably. Still, it is unclear why individuals with ASD show the opposite pattern, where BSV increases across development. This divergence may be related to a number of factors such as differences in brain maturational processes in ASD compared with TD. For example, individuals with ASD show early developmental brain overgrowth compared with TD individuals [30]. Post-mortem examinations of brains from individuals with ASD show age-related decreases in Gamma-aminobutyric (GABA) acid associated with immune and inflammation responses [31]. Such findings have led to speculation that early neurobiological differences present in ASD from early age (e.g., 2 years old) results in a cascading effect where typical brain development is affected [32]. These early developmental deviations in individuals with ASD from TD individuals may contribute to the differences in BSV found in the current study.
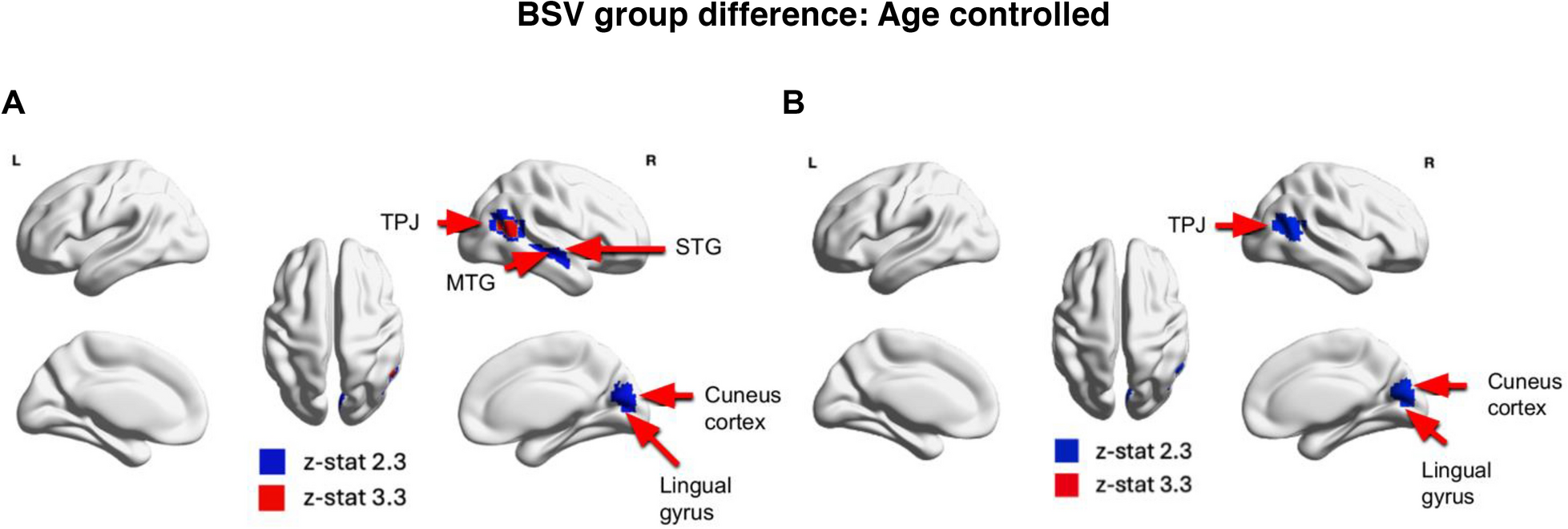
Previously, Easson and McIntosh (2019) found no group differences between ASD and TD individuals and a positive relationship between MSSD and age across both ASD and TD individuals. In contrast, we found reduced BSV in individuals with ASD compared with TD individuals when controlling for age and quadratic age in temporal and occipital cortices including the TPJ, MTG, STG, cuneus cortex, and lingual gyrus. We also demonstrate an age × group interaction where there was a positive association between rMSSD and age in the lingual gyrus, PCC, PHG, and occipital cortex in individuals with ASD while the TD group showed a negative association between rMSSD and age. There was also a quadratic age × group interaction, where individuals with ASD exhibited a slight increase in rMSSD during childhood and older adulthood, followed by a significant drop in middle adulthood in the prefrontal cortex and paracingulate gyrus, whereas TD individuals exhibit higher rMSSD in childhood and older adulthood, and lower rMSSD for middle-aged individuals.
The divergence of our results from Easson and McIntosh (2019) could arise from a number of differences between the two studies such as the sample size, age range, and the use of a multivariate (e.g., Easson and McIntosh 2019) versus univariate approach (e.g., current study). Future research should attempt to clarify how such methodological differences may influence BSV when comparing ASD and TD, when exploring how age impacts such clinical differences, and how ASD symptom severity is related to BSV. These studies underscore the complex and dynamic nature of identifying age-related changes in brain variability related to the developmental trajectories of ASD and TD individuals.
Restricted and repetitive behaviors and rMSSD
Easson and McIntosh (2019) previously identified a negative relationship between BSV and scores on the social responsiveness scale (SRS) across individuals with and without an ASD diagnosis. We investigated the relationship between rMSSD and repetitive behavior (ADI-RRB scores). While controlling for the main effects of linear and quadratic age, we found positive significant association between rMSSD and ADI-RRB scores. This pattern was evident in clusters located in the PCC, IPL, Pcun, ITG, FuG, and cuneus cortex which belong to DMN, salience and visual networks. While we found no significant linear age × ADI-RRB interaction effects, the quadratic age × ADI-RRB interaction revealed an inverted U-shaped relationship between rMSSD and age for individuals with higher repetitive behavior scores (i.e., lower rMSSD levels in both younger and older individuals, and higher rMSSD in middle-aged individuals). Those with mid-range repetitive behavior scores displayed a weak inverted U-shaped rMSSD-age association. In contrast, those with lower scores showed a U-shaped relationship between rMSSD and age (i.e., higher rMSSD levels in both younger and older individuals, and lower rMSSD in middle-aged individuals), particularly in the PrCG, PoCG, STG, and MTG region which belong to functional brain networks such as DMN, dorsal attention network (DAN), and sensorimotor network (SMN). These patterns highlight how the relationship between ASD symptom severity strongly depends on the clinical symptom, sample size, and methodological approach.
Our findings align with the neurobiological mechanisms underlying altered BSV and RRB in ASD. In ASD, an imbalance between excitation and inhibition—characterized by increased glutamate and decreased GABA—reduces the functional differentiation of brain processing systems, leading to excessive neural activity and increased noise. This heightened noise compromises the ability of the brain to process information accurately, reducing the reliability of neural representations [33, 34]. Studies in ASD rodent models have demonstrated that heightened excitatory and diminished inhibitory signaling can induce RRBs [35, 36]. Additionally, dopamine signaling plays a significant role in modulating BSV. Dopamine is integral to neuromodulation, regulating synaptic activity and modulating synaptic strength, influenced by factors such as the duration of dopamine receptor activation [37]. Depletion of dopamine in specific brain regions has been linked to increased BSV and reduced functional connectivity [38]. Altered dopamine signaling in ASD is linked to RRB-like symptoms, and studies report that dopamine agonist drugs can improve RRBs in individuals with ASD [39]. Overall, these neurobiological aspects may explain the relationship between the increased BSV and severity of RRBs in the ASD group.
The interplay between age, BSV, and RRB in ASD aligns with prior neuroimaging studies, which have reported associations between RRB and atypical functional connectivity in multiple resting-state networks across different age groups in ASD [40,41,42,43]. Abbott and colleagues reported an imbalance of cortico-striatal intrinsic functional connectivity in children and adolescents (8–17 years) with ASD. This imbalance was characterized by increased functional connectivity in limbic circuits and reduced functional connectivity in frontoparietal and motor circuits and was associated with RRB [41]. Weng and colleagues reported hypo-connectivity between PCC, mPFC, temporal lobes, and superior frontal gyrus (SFG) associated with RRB in adolescence [43]. Studies in adults with ASD revealed hyperconnectivity between PCC and the right parahippocampal gyrus [42]. Overall, these studies demonstrate that links between functional connectivity and the RRB behavioral phenotype changes from childhood to adulthood in ASD. These findings, coupled with the results of our current analysis, imply that the brain regions from DMN, DAN, SMN, and salience network exhibit age-related atypical BSV, and functional connectivity trajectories associated with RRBs in ASD. Previous studies investigating the relationship between functional connectivity and BSV have yielded mixed findings. Some report reduced functional connectivity in brain regions with increased BSV [38], while others find a positive correlation between BSV and functional connectivity [44]. Further research is required to understand the age-related dynamics of BOLD variability and its relationship with functional connectivity, especially in ASD.
The current results support the notion that there is a relationship between heightened BSV and the occurrence of RRBs; however, the direction of causality remains uncertain. It is unclear whether increased BSV precedes and influences the development of RRBs or if the observed variability patterns are a consequence of the RRBs themselves (e.g., potentially arising from ongoing efforts to manage and control them). RRBs have been observed to be less prevalent and less severe in older age groups compared with younger individuals [45, 46]. It is plausible that the increase in BSV with age among individuals with higher RRBs may serve a compensatory purpose in adulthood. These changes may compensate for age-related reductions in network complexity and integration or might indicate dysfunctional signal variability. For example, stochastic resonance (SR) may play a role in the observed increase in BSV with age, potentially reflecting a compensatory mechanism where noise is optimized to maintain neural function and processing efficiency despite age-related declines. This increase in BSV could also indicate a shift in SR dynamics, where greater external or internal noise becomes necessary to support the same level of neural integration and signal detection in older individuals, particularly those with higher levels of RRBs [47,48,49].
In contrast to individuals with elevated RRBs, we observed a reduction in rMSSD associated with lower RRB symptoms from childhood to adulthood. These findings suggest that these individuals have potentially maintained neural efficiency over time. The brains of individuals with lower RRB scores might be more capable of maintaining stable and efficient neural networks as they age, avoiding unnecessary increases in BSV that could potentially disrupt cognitive function. Future work should elucidate the relationships between RRBs and the maturation of functional brain networks across development using longitudinal datasets to enhance early risk assessment, inform developmental models of ASD pathogenesis, and provide a neurophysiological foundation for novel interventions focused on RRBs.
Limitations
There are a few limitations important to note in the present study. First, we employed a cross-sectional approach to examine age-related associations between variability and repetitive behaviors, which limits our ability to infer developmental trajectories [50]. Additionally, despite the demographic matching of groups, a cross-sectional design cannot disentangle the effects of a condition from its underlying causes. An important consideration of our cross-sectional study is the potential for misinterpreting age-related correlations, as it assumes homogeneity among younger and older individuals with the same diagnosis, disregarding the distinct characteristics associated with developmental psychopathologies that vary with age. Moreover, the inherent constraint of cross-sectional studies lies in their inability to capture individual longitudinal changes, impeding our comprehension of how the illness and related imaging measures evolve. Future research employing longitudinal designs could provide valuable insights into how the relationship between BSV and repetitive behaviors in individuals with ASD evolves.
Although our study includes participants aged 5 to 50, the number of individuals above 30 is limited due to constraints in the ABIDE dataset. Specifically, our primary analysis involved only 22 participants over 30, and the oldest participant did not exceed 30 for the ADI-RRB analysis. This skew towards childhood, adolescence, and young adulthood in our study provides an opportunity for a more detailed examination of these developmental stages. However, the limited number of older participants suggests that age x diagnosis interactions might be influenced predominantly by the few older individuals included, potentially affecting the generalizability of our findings across the entire age range. Future research should consider employing datasets with a broader and more evenly distributed age range. This would ensure a more comprehensive analysis and provide better insights into how the relationships between BSV, age, and ASD symptomatology evolve across the lifespan.
Another limitation in our study concerns potential heterogeneity within the ASD group [51]. We primarily focused on right-handed, high-functioning adult males, which could not account for all between-participant differences. Recent studies have suggested that genetic patterns may influence neural responses in autism [52, 53], and there is substantial heterogeneity among individuals with ASD [54]. To gain a more comprehensive understanding, future investigations should explore these observations in more genetically and behaviorally homogeneous subgroups of ASD individuals.
While we used the ADI-RRB scale to explore the age-related relationship between BSV and repetitive behavior in ASD, the limited availability of additional RRB measures and the smaller number of participants with these measures in the ABIDE dataset may result in an incomplete assessment of RRBs. Consequently, our findings may not capture all the nuanced relationships between BSV and RRB. Future research should consider employing a broader array of RRB assessments with a larger sample size to enhance the power and robustness of the findings. DSM-5 [55] classifies RRBs into four subtypes: (a) stereotyped or repetitive motor movements, (b) inflexible adherence to routines, (c) highly restricted, fixated interests that are abnormal in intensity or focus, and (d) hyper- or hypo-reactivity to sensory input or unusual interests in sensory aspects of the environment. Given the heterogeneity of these repetitive behaviors, there exist considerable challenges in achieving a thorough understanding and comprehensive investigation of this phenotype. Future research should aim to address these gaps and explore the associations between BSV and specific subcategories of repetitive behaviors in individuals with ASD.
Lastly, in our study, rMSSD values ranged from 0.3 to 1. The range of MSSD values depends strongly on the TR, as longer TRs allow for more changes of the BOLD signal between images. Thus, rMSSD is not capped at a maximum value of 1, but it is dependent on the TR of the fMRI scan. We previously showed that the range of MSSD for a fast TR (0.645 s) is different than the range of MSSD for a slow TR (1.4 s); additionally, the strength of linear and quadratic effects may depend on the TR [56]. Thus, the choice of TR could have influenced the linear and quadratic effects observed in our analysis. Further studies should evaluate the influence of different TR values on age-related BSV in ASD to optimize fMRI acquisition and improve assessments of brain signal variability in ASD.




留言 (0)