
記住我


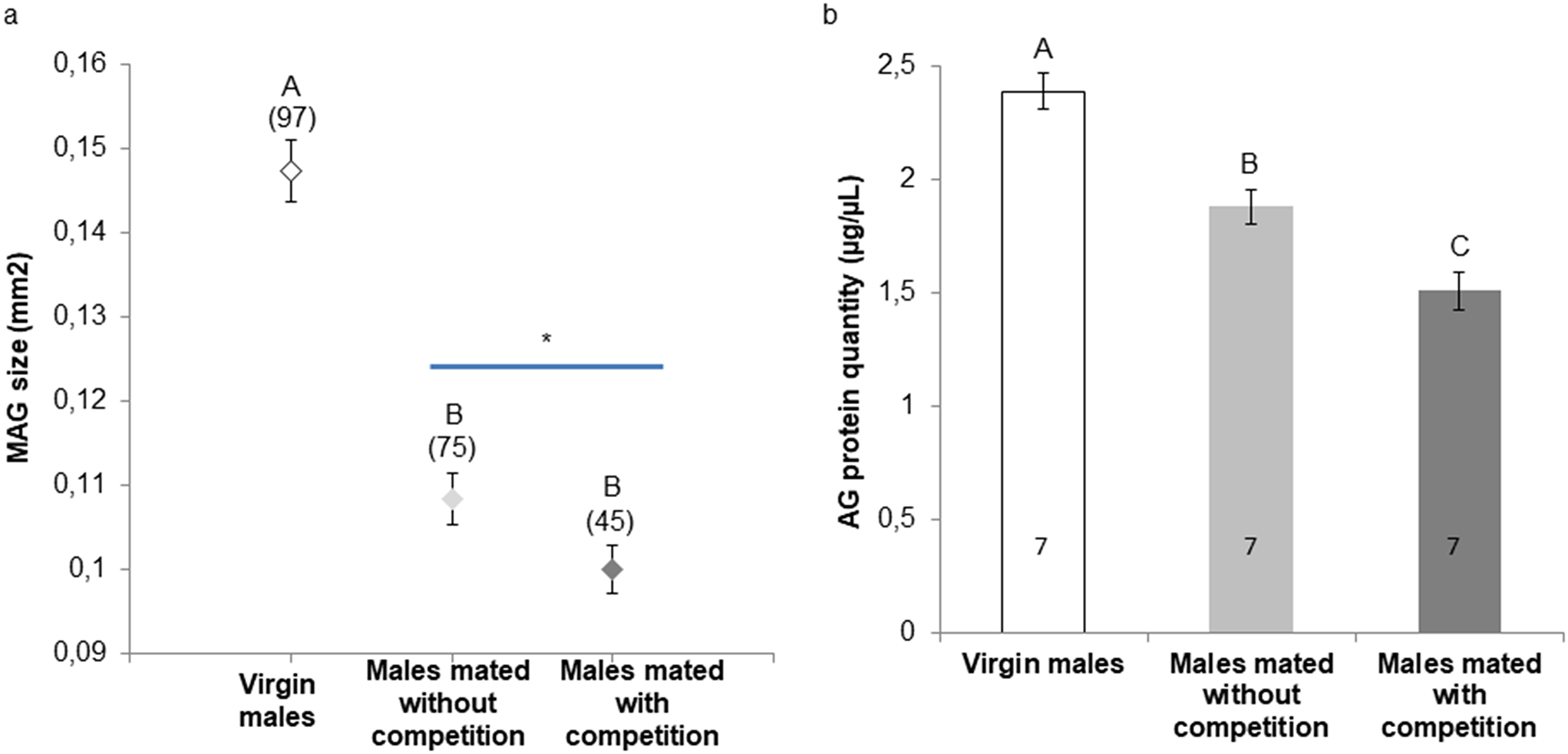

A tomato in one of the authors’ backyards fell from a vine and split open in the summer of 2021 in central Virginia. Within a day it was swarmed with various small flies and wasps. Over the next two days, the center became liquid, and many small larvae could be seen. The tomato was sectioned in half, and a glass slide was placed against the liquid center and video recorded (Fig. 1a, c). Clusters were seen to form and lasted about 12 h (Fig. 1b). The larvae in the cluster (Fig. 1b) were scooped out and placed in a processed Caltech-food fly vial in which clusters continued to form (Fig. 1c). Over the next two weeks pupae formed and hatched into adults. These were identified morphologically as 7 species of fly (Miller et al. 2017) and were placed in a new vial. Grouped larvae from this vial were tested in 2D assays and formed good clusters (Fig. 1d, Fig. s1b). These were all placed back in the vial and grown to adults. From this, breeding pairs of all species were established from which one line from each of 5 species was established. To establish inbred lines, single male–female sib pairs were crossed for 10 generations. Two species, D. tripuncta and D. algonquin, were lost after the first passage. 4 of these species are closely related drosophilids which consume rotting fruit and one, Megaselia scalaris, or Scuttle fly, is a generalist in diet.
Fig. 1
a A tomato that had fallen from a vine had ruptured and the center liquified with visible small larvae. The tomato was cut (dashed white line) and a 50 × 75 mm glass slide was placed against the liquified center. Cluster-like larval groups were seen almost immediately. These were video recorded. b Two still photos from a video shows what fulfill the definition of a cluster (Dombrovski et al. 2017). A 2 cm cube of material including the cluster in B was removed and placed in a vial. c Clusters from the tomato sample were seen in the vial. These larvae were incubated to adults and 7 species of flies and one small wasp hatched. These were allowed to reproduce and lay eggs in a separate vial. From this F1 vial, only 5 species hatched. d Samples from this mixed species F1 vial were tested for clustering in 2D assays using CS processed food. Adults hatching from this vial were separated by species into breeding pairs
Because of the potential that some members of the wild cluster could be sibs, other wild caught lines were obtained to broaden the repertoire (Fig. 2a, Fig. s1c, see methods). A number of extra D.melanogaster and one D. simulans lines, captured in different locations of the Eastern United States and established in by different labs were included in these studies. This ensures that the behaviors observed are not restricted to the initial collection location. Of 25 wild caught lines of 5 species, all cluster in vials except for D. suzukii (Fig. 2b, Fig. s2a). To quantify clustering further, 40 larvae were removed from vial clusters and placed in 2D assays using D.melanogaster CS pre-processed food as described (Dombrovski et al. 2017) (Fig. 2c). With the exception of D.suzukii, all cluster and some, like Megaselia, are almost 100% in clusters (Fig. s2b). Some lines, like the two D. simulans lines, vary significantly from each other arguing against a simple species pattern.
Fig. 2
a Each species was grouped in separate vials and produced 5 lines of 5 species. From each line, single sibs were chosen and crossed. This was done > 10 times to make inbred lines. These were combined with 19 previously established inbred lines (see materials) for further study. All together, these wild flies originate from 4 locations in the Eastern US. The relationships of the 5 species are indicated and the ones used in this study are shaded in blue. Calliphorids, which might also cluster (Scanvion et al. 2018), were not used in this study. b At least 50 adults from each of the 24 lines were placed in egg cups and 24 h old larvae were obtained. About 200 of these larvae were placed in CS-processed vials and clustering was monitored over a few days. Clusters were observed in all vials except that of D. suzukii. See Fig. s1c. 9 lines of 5 species were chosen for further study. c Larvae were allowed to cluster in CS-processed vials as described (Dombrovski et al. 2017). 40 larvae were removed and placed in a 2D apparatus with CS-processed food. The percentage of larvae in clusters at 4,5 and 6 h was averaged. All larvae cluster with the exception of D. suzukii. Almost all larvae of Megaselia larvae are in clusters. Points indicate average values and error bars represent the standard error. The number of 2D samples used is indicated on the X-axis. Statistical probabilities were calculated by ANOVA, after normality tests, followed by Tukey’s method. The probabilities compared to CS are indicated with significance marked by P < 0.01 ** and P < 0.001 ***. A T-test was performed between simR and sim2 and is significant P < 0.0001. d Clustering residing times of transplanted larvae. Third instar larvae were removed from crowded vials with clusters and labeled with food coloring. Single larvae were placed over clusters of host larvae and the residence time was measured as described (Dombrovski et al. 2017, 2019). With the exception of M1 and suz, all spend 20-60’ in clusters. suz larvae do not enter CS clusters but data is included on the graph for clarity. There is also asymmetry in many transplantations, like M1- > CS/CS- > M1, DmR- > CS/CS- > DmR and sim2- > CS/CS- > sim2. Points indicate average values and error bars represent the standard error. The number of 2D samples used is indicated on the X-axis. Statistical probabilities were calculated by ANOVA, after normality test, followed by Tukey’s method. The probabilities compared to CS- > CS are indicated with significance marked by P < 0.01 **, P < 0.001 *** and P < 0.0001 ****. A T-test was performed between the 3 reciprocal pairs. e Inter-larval synchrony time was measured for transplanted larvae and compared to residing time in clusters. When measured for the different wild types, synchrony explains some of the variance in cluster residence time. However, some cases of long residence time, like sim2- > CS, seems to happen without a high degree of synchrony. f To test for the potential effects of species-specific food processing, food from 3 species was used to host CS 2D clusters. Plotted is the proportion of CS larvae clustering and resultant wing size of emerging adults from matched vials. Compared to food processed by CS, that processed by Megaselia, Zaprionus and simulans supports CS is clustering and gives healthy adult wing sizes. Only Zaprionus food gives lower wing sizes for CS. The data points are the averages of the indicated number of samples. The error bars indicate standard deviation for wing size and standard error for clustering. Statistics were calculated by normality tests followed by ANOVA and Tukey’s test. The probabilities are indicated and * is P < 0.05
To test for the idea of co-clustering, transplantation experiments were conducted of larvae from species shown in (Fig. 2) into D.melanogaster CS clusters. In these experiments, food-coloring tagged larvae were placed into pre-established CS clusters as described (Dombrovski et al. 2017). CS host clusters were established from clusters in vials and on CS-preprocessed food. For intruders, larvae were removed from clusters in vials, placed in food coloring for 30’ and placed over CS clusters. The residing time was measured as how many minutes an intruder spent in a host CS cluster (Fig. s2c, d, e, f). Times vary greatly and do not necessarily correlate with single species clustering measures. For instance, sim2 does not cluster well with itself but resides in CS clusters as well as CS larvae themselves (Fig. 2c, d). High resolution movies were made of transplants and the inter-larval delay time in movement was used to measure the degree of synchronization (Dombrovski et al. 2017) (Fig. s2f, g). Plotting the synchrony time versus residence time in clusters reveals a relationship (Fig. 2e). The shorter the synchrony time, which means more alignment of the transplanted larva to the hosts, the longer the intruder resides.
For all measures of these wild caught flies, CS processed food was used. The clustering parameters for this food have been well established (Liao et al. 2024). To examine the potential role of species-specific processing, food was processed as described (Dombrovski et al. 2017) but using simR, zap or meg as the feeders. CS 2D clustering was examined on all three food types and compared to that processed by CS. In addition, 40 L2 CS larvae were loaded into a vial with food processed by one of the 3 wild types, and wing size from emerging adults were measured (Fig. 2f). None of the clustering measures were significantly different from that on CS food (Fig. 2f), although simR is trending to lower clustering. For wing size, zap food produces slightly lower wing size but not simR or meg. This means that while food processing can have an effect on clustering of other species, the effect is small.
To examine the effects of larval mixed foraging on adult fitness, female wing size was used, as described (Dombrovski et al. 2020). Wing size is a good indicator of body size which relates to fecundity (Gilchrist and Partridge 1999). For each line, 10 and 40 L2 larvae were loaded into separate vials based on the finding that wing size is larger at 40 compared to 10 animals per vial (Liao et al. 2024). An increase in wing size going from 10 to 40 indicates an advantage with greater numbers (Liao et al. 2024).Larvae were then mixed with the benchmark lab ‘P’ strain which clusters well with CS (Liao et al. 2024). This strain is a lab-based transgenic and inbred line which has been used as an alternative to CS (Liao et al. 2024). These were mixed at 10:30 and 30:10 ratios such that there were 40 larvae per vial. Wing size of the test line and P was established for each experiment (Fig. 3). This allows an estimation of the value of clustering, the value of mixing with the lab P strain and the effects on P. For comparison, data from (Liao et al. 2024) are replotted in this format in Fig. 3a. CS larvae produced bigger wings when in groups of 40, either alone or mixed with P and P is equally advantaged by this arrangement (Fig. 3a). This is schematized in Fig. 3j where arrow size indicates gain (large arrow) or neutral/loss (small arrow). For the 3 D.melanogaster lines, M1, DmR and C8, each behaves in a different manner (3a, b, c, d). In general, CS and DmR show mutualistic gain with P while M1 and C8 do not.
Fig. 3
Larva fitness was measured by female adult wing size (Y axis). Second instar larvae were placed in a vial, as 10x, 40x, 10 × and 30 × wild type D.melanogaster host ‘P, and 30 × with 10xP. The effects of intra-specific clustering can be seen in any change from 10 × to 40x. The effects of heterospecific blends can be measured by comparing 40 × with either 10–30 or 30–10 blends with P. The effects on P are summarized in the right graph for each panel. The blue line represents the average 40xP wing size. The individual wing sizes are shown with the red line marking the mean. Statistical significance was measured by ANOVA post a normality test followed by Tukey’s method. The upper of the two probabilities compares to 40 × and the lower to 10 × of the same kind of larvae. P < 0.05 *, P < 0.01 **, P < 0.001 *** and P < 0.0001 ****. For P host measurements, the probability is compared to 40xP (Liao et al. 2024). a CS: data is replotted and analyzed from before (Liao et al. 2024). CS gains more from P than P from CS. b DmR neither gains nor loses to P. c M1 loses fitness to P and P remains the same. d C8 and P lose fitness in blends. e simR and P lose fitness in some blends. f sim2 loses in self clustering, and also loses in blends while P gains. g suz loses in blends while P gains. h zap loses in blends while P remains neutral. i meg and P lose fitness in some blends. j Summary of fitness gains or losses in blends with P for each of the 9 lines tested. An arrow pointing to the P host indicates a gain in fitness of P and an arrow to the test line a gain of that line. The self referential arrows indicate gains of self clusters. The large arrows indicate gains while the small arrows either a loss or neutral effect. There is not a species pattern to the gains or losses
Similarly for D.simulans, one line shows a mutualistic gain with P, (sim2, Fig. 3f) while the other does not (simR, Fig. 3e). All three of the other species, M. scalaris, D.suzukii and Z. indianus show mixed interactions with P. P gains from all three in some of the mixes.
To examine for broader relationships, wing size and clustering data from. s 2 and 3 were compared. To examine the role of self-clustering, the wing effects of clustering (wing size at 40 compared to wing size at 10, first two columns in Fig. 3) were compared to how much clustering each line does (Fig. 2c). No relationship is seen (Fig. 4a). The relationship between time spent by an intruder in a cluster (Fig. 2d) and gain from that cluster (wing size 10/30P divided by 40, or third column in Fig. 3 divided by second) gives a strong relationship. CS and sim2 spend most time in P and gain the most while suz spends least time and gains the least. Interestingly, plotting the same x axis against the gain for the host P (Fig. 4c, P wings in 10/30P divided by 40xP alone) also shows a relationship. If the intruder gains, so does the host. This is further clarified in Fig. 4d by plotting the gain of the intruder over the gain of the host. If the intruder gains, so does the host.
Fig. 4
Clustering and fitness data from wild caught flies were assembled and correlations examined. Four selected lines are indicated for each graph. a The relationship between the amount of clustering any line conducted with itself was compared to the gain from that behavior. The gain from mono-species clustering was measured by wing size for 40 × larvae in a vial divided by that from only 10 larvae. This was compared to proportion of larvae that cluster in 2D. b The gain in fitness of an intruder in P was measured by wing size at 10/30 blend compared to 40 alone. This is plotted against time spent by an intruder in a 2D P cluster. c The same relationship in 4B is now plotted but now as the gain of the host versus the time spent by the intruder in that host cluster. d Gain of intruder is plotted against the gain of the host
留言 (0)