
記住我
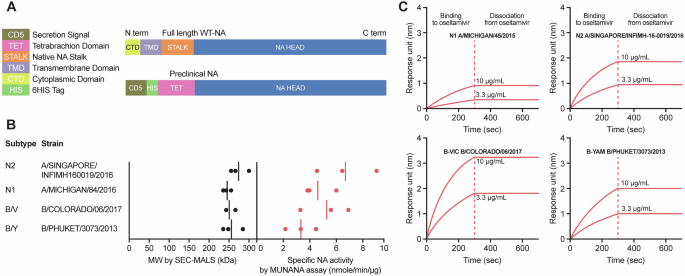
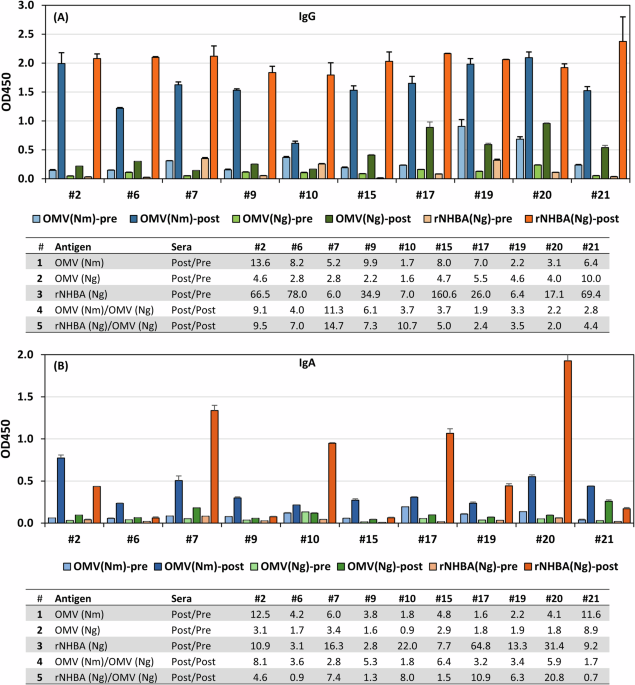
Previous work has shown that the OMV (Nm strain NZ98/254) component of 4CMenB generates serum antibodies that cross-react with proteins and LOS in Ng OMV20,21,22. To assess the cross reactivities of the 4CMenB vaccine-induced antibodies to gonococcal antigens, OMVs were isolated from the spent culture supernatants of Nm vaccine strain NZ98/254, the widely used laboratory gonococcal strain 1291 and a recent gonococcal urethral clinical isolate CNG20. The gonococcal strains express the two most common NHBA variants that account for >60% of the Ng genome records (n = 16,315) in the PubMLST database. In addition, recombinant NHBANg (rNHBANg) proteins expressed by 1291 (NHBA_542) and CNG20 (NHBA_475) were cloned and purified from E. coli as mature proteins without the signal peptide. These two most common gonococcal NHBANg proteins are highly similar (99.3% identity), with only three residue variations (A390V, G414S, and S434G) at the C-terminal domain. In preliminary analyses of IgG and IgA recognition of the two gonococcal OMVs and two rNHBANg proteins, no strain-dependent differences in antibody responses of 4CMenB sera between two gonococcal OMVs and between two rNHBANg antigens were observed, thus the OMVCNG20 and rNHBA1291 were selected as the representative of OMVNg and rNHBANg, respectively. The OMVs from the vaccine strain NZ98/254 was used as a reference (OMVNm) for comparisons to the gonococcal antigens. We subsequently probed the IgG and IgA responses to OMVNm, OMVNg, and rNHBANg by ELISA with 10 matched sets of pre- and post-4CMenB sera. The post-sera were obtained 1 month after the third dose of 4CMenB23. All ten post-4CMenB sera showed increases in IgG responses to OMVNm (1.7–13.6-fold) when compared to the corresponding pre-sera (Fig. 1A); the post serum #10 had the lowest IgG response (1.7-fold). Two pre-sera (#19 and #20) showed higher baseline IgG levels than the other pre-sera, implying potential preexisting antibodies. The IgG levels detected by OMVNg paralleled those for OMVNm with 1.6–10.0-fold increases post 4CMenB immunization; however, the recognition of OMVNg was lower compared to OMVNm, both in baseline and post vaccination sera (1.9–11.3-fold, Fig. 1A Table). As NHBA is the only recombinant protein of the 4CMenB vaccine that is surface expressed in Ng, we assessed the antibodies specifically targeting NHBA using purified rNHBANg as the capturing antigen in ELISAs. All post 4CMenB sera showed very strong IgG reactivity to the rNHBANg protein and the IgG of post 4CMenB sera compared to the pre-sera increased 6.0-160.6-fold, which was not surprising considering there is 50 μg recombinant NHBA-GNA1030 fusion protein in a 0.5-mL dose of 4CMenB vaccine. The markedly higher IgG signals captured by rNHBANg relative to those captured by OMVNg (2.0–14.7-fold, Fig. 1A Table) indicated that 4CMenB vaccination induced a strong anti-NHBANm response that cross-react with NHBANg. However, the amount of NHBANg present in the gonococcal OMVs based on the lower signal intensity appeared limited. These signal differences were not caused by different variants present in the OMVNg and rNHBANg coating antigens as IgG recognitions of NHBA variants expressed by 1291 and CNG20 are near identical (Supplementary Fig. 1). In addition, the limited amount of NHBA in gonococcal OMVs preparations is supported by Western blots (Fig. 2, lane 3). Weak or no signal was seen at the position of NHBANg in OMVNg samples despite there were 5-fold higher protein loadings of OMVNg preparations relative to the OMVNm samples, which showed NHBANm signals.
Fig. 1: 4CMenB immunization generated individual variability in IgG and IgA responses to OMVs and rNHBA.
ELISA plates coated with purified Nm NZ98/254 OMVs (blue), Ng CNG20 OMVs (green) or rNHBA1291 (orange) were probed with pre- and post- sera followed by anti-human IgG-HRP (A) or anti-human IgA-HRP (B). Tables show the IgG or IgA fold increases, comparing post/pre of OMV (Nm), OMV (Ng), and rNHBA (Ng), or comparing post/post of OMV (Nm)/OMV (Ng) or rNHBA (Ng)/OMV (Ng).
Fig. 2: Western blots of IgG responses of pre- and post- 4CMenB sera to gonococcal OMV and rNHBA.
Western blots of Nm OMVNZ98/254 (2 μg, lane 2), Ng OMVCNG20 (10 μg, lane 3) and purified recombinant NHBA1291 (500 ng, lane 4) probed with 10 individual pre- (top panels) and post-4CMenB (bottom panels) sera followed by anti-human IgG-HRP. Lane 1, MW markers. The positions of NHBA, FHbp, and GNA2091 are marked by arrows. Relative to the NHBA of the Nm NZ98/254 strain, there is a ~60-aa deletion in the gonococcal NHBA. The reactive lower MW bands in lane 4 in several blots are likely degraded NHBA protein. The position of the 75-KDa marker is indicated with a black arrow in each blot.
We also evaluated the serum IgA responses by ELISA and observed considerable individual variability toward OMV antigens among ten paired sera (Fig. 1B). Increases of post vs pre sera were 1.6–12.5 fold and 0.9–8.9 fold for OMVNm and OMVNg, respectively (Fig. 1B Table). The IgA responses were, as expected, higher against OMVNm than OMVNg (1.7–8.1-fold, Fig. 1B Table). Relative to the IgA responses to OMVNg, the IgA reactivity to rNHBANg was higher for most post-sera (0.7–20.8-fold, Fig. 1B Table), which is in contrast to the consistently high IgG reactivity to rNHBANg. Immunization-induced IgA responses to rNHBANg were robust in over half of the post vaccination sera (#2, 7, 10, 17, 19, and 20), each with greater than a 10-fold increase in post vaccination recognition (10.9–64.8-fold), but were lower for the other sera.
To define the OMV proteins that were reactive with 4CMenB antibodies and confirm the specific recognition of gonococcal NHBA, we performed Western blots using both pre- and post- 4CMenB sera to probe IgG (Fig. 2) and IgA (Fig. 3) responses. As the signal of OMVNg were lower than that of OMVNm in ELISA, a 5-fold higher amount of OMVNg relative to OMVNm was loaded to obtain similar signal intensities between two OMV samples and enable the detection of weak reactive bands in OMVNg without over-exposure of signals from OMVNm. Pre-existing antibodies were detected in most of the pre-sera (Top panels). These antibodies recognized proteins usually <75 kD in OMVNm (lane 2) and/or OMVNg (lane 3), but no signals were detected for rNHBANg in pre-sera (lane 4). IgG signals for OMVNm were detected in pre-sera of #7, #10, and #20 and there were IgA signals in pre-serum #17. For OMVNg, pre-sera of #7, #17, and #20 showed IgG bands, and IgA bands were in pre sera of #2, #9, #17, and #21. For the post sera (Bottom panels), recognition of multiple Nm and Ng OMPs and rNHBANg was observed. The dominant IgG responses appeared to target the three recombinant antigens of the 4CMenB vaccine (GNA2091, FHbp, and NHBA) in OMVNm (lane 2). For OMVNg, the signals of GNA2091, which is localized at the periplasmic side of the outer membrane18 and in the vaccine as the C-terminal protein of the FHbp-GNA2091 fusion, were the highest for all post sera. As expected, there was no signal for FHbp in the OMVNg samples (lane 3). The position of the rNHBANg in Western blots was lower than that of meningococcal NHBA, reflecting a 60-amino acid (aa) deletion in gonococcal NHBA described below. The NHBA_2 variant in the 4CMenB vaccine has ~85% protein sequence similarity to gonococcal NHBA_542 and NHBA_475 variants and the two NHBANg proteins are highly similar (99.3% identity) (Fig. 5A). IgG antibodies in post sera clearly reacted with gonococcal rNHBANg (lane 4), however, no, or very weak reactivity was detected at the corresponding NHBA position in the OMVNg samples (lane 3 vs. lane 4), even though there was a 5-fold higher protein loadings of OMVNg relative to the OMVNm samples with detectable NHBANm bands. This result was consistent with the ELISA data and again suggested that NHBA expression on the gonococcal outer membrane was limited.
Fig. 3: Western blots of IgA responses of pre- and post- 4CMenB sera to gonococcal OMV and rNHBA.
Western blots of Nm OMVNZ98/254 (2 μg, lane 2), Ng OMVCNG20 (10 μg, lane 3) and purified recombinant NHBA1291 (500 ng, lane 4) probed with 10 individual pre- (Top panels) and post-4CMenB (bottom panels) sera followed by anti-human IgA-HRP. Lane 1, MW markers. The reactive lower MW bands in lane 4 in several blots are likely degraded NHBA proteins. The position of the 75-KDa marker is labeled with a black arrow in each blot.
In contrast to the consistent IgG recognition of rNHBANg by all ten post 4CMenB sera, variable recognition of rNHBANg by IgA antibodies was observed (Fig. 3), correlating with the ELISA data shown in Fig. 1B. For example, strong rNHBANg signals were seen with post sera #7, #17 and #20, whereas post sera #6 and #15 yielded faint signals. In addition, the IgA signals to GNA2091 and FHbp (in OMVNm) were also highly variable in comparison to the IgG responses. No correlation of IgG and IgA signal intensities was seen between GNA2091 and FHbp in OMVNm. For example, the IgG signal intensity of FHbp in OMVNm was greater than that of GNA2091 for post serum #19, but the reverse was true for the IgA responses of post sera #19.
Western blots of nhba mutants confirmed the presence of NHBA antibodiesTo confirm the reactive band near the 75-kD molecular weight (MW) marker was indeed NHBA, an nhba::aphA3 deletion-insertion mutation was created in two meningococcal strains, NZ98/254 and CNM3, and two gonococcal strains, 1291 and CNG20. CNM3 is a representative isolate of the N. meningitidis urethritis clade adapted to urogenital tract. Gonococcal strains 1291 and CNG20 express the two most common gonococcal NHBA protein variants (NHBA_542, 31.8%, and NHBA_475, 28.7%). OMVs prepared from WT and the nhba mutants were probed with post-sera #19 by Western blot (Fig. 4). In Nm, three dominant signals were observed, corresponding to the recombinant antigens present in the vaccine, GNA2091, FHbp and NHBA. As anticipated, the FHbp signals were absent in both gonococcal strains, indicating that the gonococcal homolog, Gfhp, is not present in OMV. The reactive proteins near the 75-kD MW marker in all WT strains were absent in all nhba mutants (Fig. 4), indicating the signal was NHBA. The NHBA bands of both gonococcal strains and CNM3 migrated faster than NHBA of the vaccine strain NZ98/254 due to the N-terminal deletion of ~60 residues (see Fig. 5A).
Fig. 4: Western blot of purified OMVs from WT gonococcal and meningococcal strains and corresponding nhba mutants.
Aliquots of OMV preparations (15 μg total protein content) from each strain, except the WT NZ98/254 at 2.5 μg, were probed with post 4CMenB sera #19 and anti-human IgG-HRP. NHBA proteins in WT of Ng (1291 and CNG20) and Nm (CNM3 and NZ98/254) strains, but not in nhba mutants (NHBA -), were recognized by post sera #19. FHbp is found in Nm OMV, but not in Ng OMVs. GNA2091 is fused to FHbp in 4CMenB to enhance immunogenicity and stability and is also recognized by post sera #19. The locations of GNA2091 (22 kD), FHbp (27 kD) and NHBA are marked. The intensely reactive 27-kD band absent in Ng determines the position of FHbp. The ~22-kD band is assigned as GNA2091 based on the notation in Western blots of ref. 21. Nm NHBA_2 (NZ98/254) runs above the 75-kDa MW marker, while NHBAs of Ng strains and CNM3 migrated faster than the 75-kDa marker as anticipated based on the ~60-aa deletion.
Fig. 5: Sequence comparison of NHBA variants.
A Protein sequence alignment of meningococcal and gonococcal NHBA variants. Gonococcal strains 1291 (NHBA_542) and CNG20 (NHBA_475) express the Ng dominant variants. Both variants, differing by three amino acids, contain identical ~60-aa deletions at the N-terminal region when compared to NHBA _2 of Nm strain NZ98/254. The NmUC isolate CNM3 expresses the NHBA_20 variant with a similar deletion. NHBA_3 and NHBA_17 variants examined in previous NHBA studies25 are also included. The variant ID is included in the parenthesis following each strain. Amino acids different from that of NHBA_2 are colored. The amino acids of the arginine rich motif responsible for heparin binding are in red text in the NHBA_2 sequence. The meningococcal NalP generated the C2 fragment of NHBA, while human lactoferrin (hLf)15, C3-convertase (C3bBb)33, saliva kallikrein 1 (hK1), and plasma Kallikrein (PKa)34 cleave at the same site, releasing the C1 fragment of NHBA. The % sequence similarity and identity are shown in the table. B The prevalence of top 10 gonococcal NHBA variants.
Analysis of meningococcal and gonococcal NHBA proteinsA search of the PubMLST database (performed 3/5/2024) identified 16,305 Ng records having NHBA (NEIS2109). Among these, 46 NHBA peptide variants (426-429 residues in length) were found in 87.7% of these isolates, while the rest were without an assigned variant number. Fourteen NHBA variants represented 97.7% of Ng isolates, of which nine variants have >2% prevalence and together accounted for 81.1%. NHBA_542 (strain 1291, 31.8%) and NHBA_475 (strain CNG20, 28.7%) are present in > 60% gonococci (Fig. 5B). There were only 1-5 residue differences among the top 10 NHBANg variants. The NHBA_2 in the 4CMenB vaccine strain NZ98/254 was not found in Ng, although there was~80% protein similarity between NHBA_2 and the two major gonococcal NHBAs. Of note, NHBA_20 of the meningococcal urethritis clade (NmUC) strain CNM324 had ~90% similarity to the gonococcal homologs (Fig. 5). NHBA_20 in NmUC and NHBA_17 found in Nm strain 2996, which was examined in previous NHBA related vaccine studies25, shared the deletion of ~60 residues in the gonococcal NHBA variants. Processing of NHBA by bacterial (NalP) and human proteases is centered around the highly conserved Arg-rich motif. Curiously, a second deletion near the arginine rich motif of the gonococcal NHBA variants, but not in Nm, would appear to eliminate the NalP cleavage site (Fig. 5A).
Serum bactericidal activities of post 4CMenB sera against N. gonorrhoeaeComplement-fixing antibodies directed against three recombinant proteins and OMV antigens (both OMPs and LOS) in the vaccine mediate 4CMenB-induced serum bactericidal activity (SBA). SBA has been established as a surrogate for protective immunity against Nm26. While the role of SBA in protection against gonococcal infections is not clear, SBA continues to be a useful assay for evaluating functional antibody-mediated vaccine responses. Thus, we investigated the activity of 4CMenB-induced bactericidal antibodies against two gonococcal strains, expressing the most commonly found NHBA variants, and compared to the Nm vaccine strain NZ98/254. Initially, we tested three individual paired pre- and post-4CMenB sera against Nm strain NZ98/254 (Supplementary Fig. 2). As expected, the post sera showed stronger SBA relative to the pre-sera from the same individuals, indicating vaccine-induced systematic antibody response. Post serum #10 displayed lowest SBA among these three post sera against NZ98/254—22% survival at 10% sera. This correlated with the lowest IgG response to OMVNm as detected by ELISA (Fig. 1A). At the highest concentration tested (10%), <10% survival was observed for post sera #7 and #17 against the vaccine strain NZ98/254. However, the same 10% post sera #7 showed less activity against gonococcal strain 1291 than post sera #17 (~87% vs. 66% survival, respectively). Therefore, additional post sera were examined against Ng strain 1291 at 10% (v/v) concentration. Low bactericidal activities were also observed for post-sera #6, #9, and #15 (84-87% survival). Three post sera, #2, #17, and #19, with higher SBA (~65% survival) are shown in Fig. 6. All three post sera exhibited higher SBA than the corresponding paired pre-sera with 2-5-fold rise in bactericidal activity post 4CMenB immunization. The lower bactericidal activity toward Ng compared to Nm correlated with the overall lower antibody recognition of gonococcal OMV antigens detected by ELISA (Fig. 1).
Fig. 6: 4CMenB vaccination induced serum bactericidal activity against Ng.
Percent survival of paired pre- (light color) and post-sera (dark color) tested at 10% (v/v) against gonococcal strain 1291. Data are presented as the mean and standard deviations with individual data points of biologically independent experiments shown as black dots. Statistically significant differences between paired pre and post-sera were determined by two-tailed unequal variance Welch t tests (**, p < 0.01; ***, p < 0.001).
Serum bactericidal activity of post 4CMenB sera targeting NHBAThe weaker SBA against Ng relative to Nm was not surprising considering that major antigens in the 4CMenB vaccine were either not present (NadA and PorA) or not expressed on the surface of the gonococcus (the gonococcal homolog of FHbp, Ghfp)19. The remaining major recombinant antigen in 4CMenB is NHBA. We evaluated the importance of 4CMenB-induced NHBA-specific antibodies by competitive SBA experiments with post sera #2, #17 and #19, which showed appreciable bactericidal activities against Ng (Fig. 6). Post sera were pre-incubated with increasing amounts of rNHBA proteins to compete out NHBA-specific antibodies and % killing in the absence of rNHBA was set as 100% to evaluate the effects of titrating out NHBA-specific antibodies. Initially, post serum #17 at 10% (v/v) was tested against the Nm vaccine strain NZ98/254 and there was near complete killing with no significant titration with up to 100 ng rNHBA (black line, Supplementary Fig. 3). Thus, for Nm strain NZ98/254 the serum concentration was lowered to 5% to enhance the sensitivity of rNHBA titration, whereas 10% sera was kept for Ng strains. Both the meningococcal NHBA_2 and gonococcal NHBA_542 proteins were first assayed and all titrations were saturated at 50 ng rNHBA under the experimental conditions (Supplementary Fig. 3). There was no difference in the titration of meningococcal NHBA_2 or gonococcal NHBA_542 against Ng strain 1291 or Nm strain NZ98/254 (Supplementary Fig. 3), implying common epitope(s) shared between NHBA variants sequestered NHBA-specific antibodies. Subsequently, all three post-sera, #2, #17, and #19, were examined by competitive SBA with Nm strain NZ98/254 and Ng strain 1291 using the corresponding rNHBA variant expressed by the strain (Fig. 7A). For the Nm strain NZ98/254, only ~10-20% of bactericidal activities of post-sera #2 and #17 were titrated away, while there was no reduction seen with post-sera #19 up to 50 ng rNHBA. Thus, the removal of anti-NHBA antibodies had limited effect on killing of Nm strain NZ98/254 because bactericidal antibodies targeted other antigens, such as FHbp, were present in the vaccine strain, likely at a higher level. However, while post-sera, #2, #17, and #19, only yielded ~30–40% killing against Ng strain 1291 at 10% serum concentration (Fig. 6), ~80% of this killing was titrated away by the presence of rNHBA (Fig. 7A). To independently support this observation, we tested one additional gonococcal strain CNG20 with competitive SBA assays. As shown in Fig. 7B, the titration profiles of all three post-sera against CNG20 were similar to each other and analogous to those for Ng strain 1291. Thus, NHBA-specific antibodies contributed to a majority of bactericidal activities against gonococci.
Fig. 7: Competitive serum bactericidal activity (SBA) assays with NHBA proteins.
Competitive SBA assays of post 4CMenB sera, #2, #17, and #19 against gonococcal strains 1291 (A) and CNG20 (B) at 10% (v/v) and against Nm strain NZ98/254 at 5% (v/v). The rNHBA protein used in titration is the intrinsic variant of each target strain. Reactions with post 4CMenB sera only without adding rNHBA are set as 100% for normalization. Each titration was repeated three times except sera #17 and #19, which were two times due to a limited amount of post sera available. The percentages of killing for post sera #2, #17, and #19 before normalization against strain 1291 are 42.4 ± 9.9, 37.4 ± 3.8 and 38.8 ± 0.4; for strain CNG20 are 41.0 ± 2.6, 39.0 ± 2.1, and 37.4 ± 4.2; for strain NZ98/254 are 73.1 ± 2.0, 88.8 ± 4.1, and 100.0 ± 0.0. Data of Nm strain NZ98/254 are included in both (A, B) for comparison to Ng.
Removing the antigen targeted by the key bactericidal antibodies was expected to impact strain susceptibility to SBA. Thus, Δnhba mutants in both the Ng 1291 and Nm NZ98/254 backgrounds were evaluated for the contribution of NHBA-specific antibodies to overall SBA. No significant difference in serum sensitivity was observed between WT and the Δnhba mutant of the NZ98/254 strain (Fig. 8A, red lines), whereas greater than 90% of the Ng 1291Δnhba mutant survived in the presence of 10% post-serum #17 as compared to 70% survival of the WT 1291. This effect was lost at lower serum concentrations. These data were consistent with the competitive SBA assays (Fig. 7), in which rNHBA titrated out most bactericidal activity against gonococci. The enhanced serum resistance of the nhba mutant was further confirmed with two other post-sera, #2 and #19, also tested at 10% (Fig. 8B). Finally, we have also compared another Ng strain CNG20 and the CNG20Δnhba mutant with 10% post-4CMenB serum #2. The CNG20Δnhba mutant indeed survived better relative to the parental WT CNG20 (75.3 ± 5.1% and 53.6 ± 6.3% survival for mutant and WT, respectively, p < 0.01). Thus, the enhanced serum resistance to post-4CMenB sera upon an Δnhba mutation was independently reproduced in two gonococcal strains.
Fig. 8: The Δnhba mutation in N. gonorrhoeae strain 1291, but not N. meningitidis strain NZ98/254, enhanced survival in serum bactericidal activity assays.
A SBA assays of post 4CMenB sera #17 against the WT (solid lines) and the Δnhba mutant (dotted lines) of Ng strain 1291 (blue) and Nm strain NZ98/254 (red) in the presence of 10% IgG- and IgM-depleted normal human serum pool as the complement source. Two-way ANOVA with Tukey’s HSD test was used for statistical analyses (*, p < 0.05; ****, p < 0.0001). B Three post-4CMenB sera were assayed at 10% (v/v) against the WT (solid) and the Δnhba mutant (stripped) of Ng strain 1291. Data are presented as the mean and standard deviations with individual data points of biologically independent experiments shown as black dots. Statistical significance was determined by two-tailed unequal variance Welch t tests (***, p < 0.001; ****, p < 0.0001).
Evidence of synergy in bactericidal activities between anti-NHBA and anti-LOS antibodies against N. gonorrhoeaeWe recently found that removing anti-LOS antibodies in post 4CMenB sera significantly reduced the SBA against Ng strain 129120. As shown in Fig. 7, titration with rNHBA also removed a significant portion of SBA against strains 1291 and CNG20. These two sets of data suggested that anti-NHBA and anti-LOS antibodies may act cooperatively in the SBA assay27. To address, competitive SBA assays [again using 10% (v/v) post 4CBMenB sera] were conducted with strain 1291 in the absence or with pre-incubation of the two antigens individually or in combination. Lower amounts of competing antigens (e.g., 1 ng rNHBANg alone, 1 ng purified L3 LOS alone, or 1 ng rNHBANg + 1 ng L3 LOS) were used to assess synergy. The percentage killing of the control reaction with 10% post-sera #2 was set as 100% to define the titration effect of pre-incubation with exogenous antigens. As shown in Fig. 9, pre-incubation of sera with 1 ng of either rNHBANg alone or L3 LOS alone reduced ~30% of the bactericidal activity of the serum. However, absorption with both antigens titrated away ~90% of the SBA.
Fig. 9: Evidence of synergy of anti-NHBA and anti-LOS antibodies in post 4CMenB sera in serum bactericidal activity (SBA) against N. gonorrhoeae.
The competitive SBA assays were conducted with strain 1291 in the presence of 10% IgG & IgM depleted human sera (complement source) and 10% post-sera #2 (heat-inactivated) with a 30-min pre-incubation of the following antigens: (1) PBMI control, (2) 1 ng rNHBA1291 alone, (3) 1 ng purified L3 LOS alone, and (4) 1 ng rNHBA1291 + 1 ng L3 LOS. The reaction with complement only was used for normalization to calculate % killing under each condition. The % killing of the PBMI control (10% post sera only; 38.6 ± 0.2) was set as 100% to determine the titration effect of pre-incubation with exogenous antigens. Data are the mean and standard deviations with individual data points of biologically independent experiments shown as black dots. One-way ANOVA with Tukey’s multiple comparison test for pairwise comparisons across each of the variables was applied for statistical analysis (****, p < 0.0001).
留言 (0)