
記住我







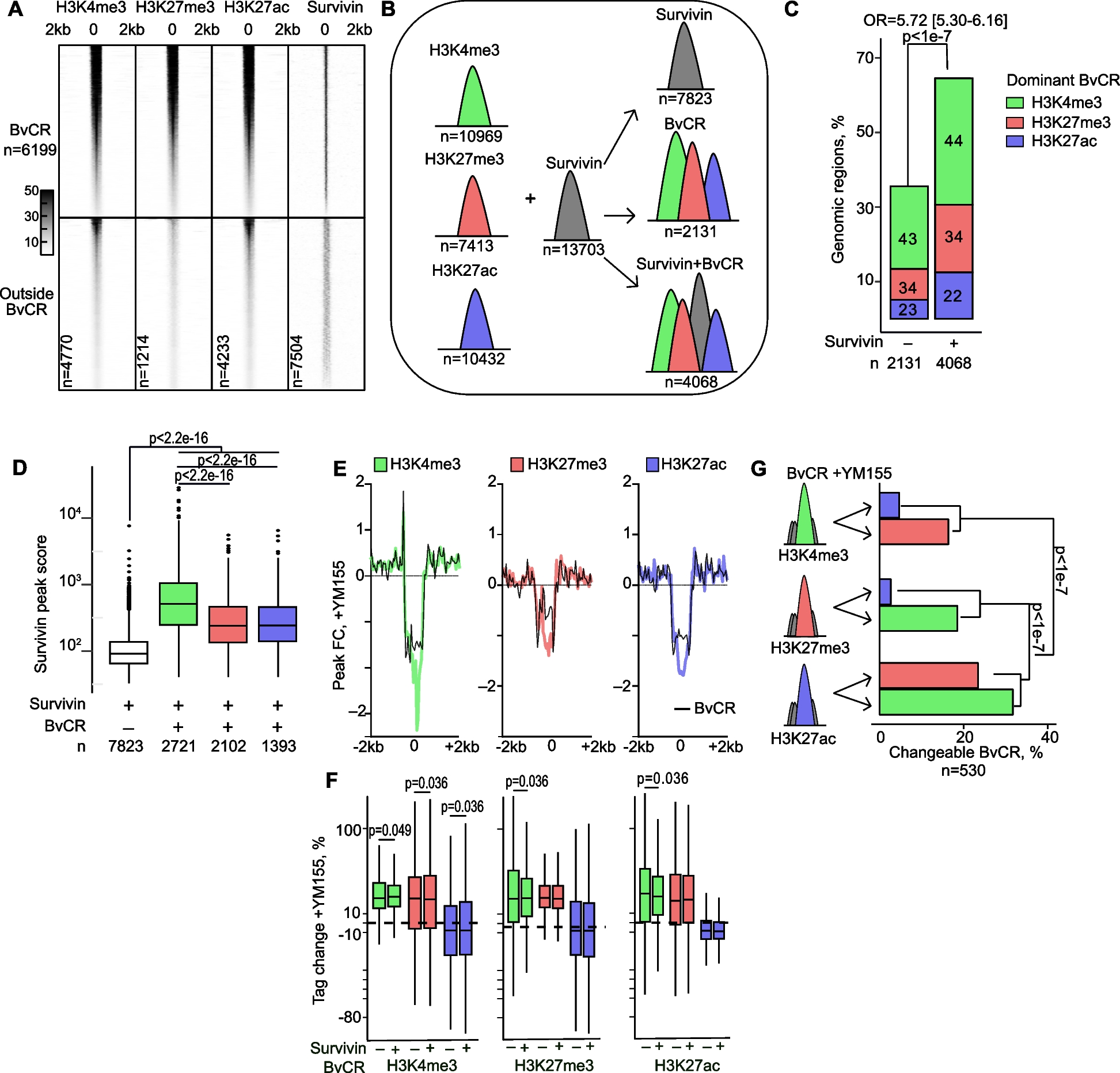


The mammalian skin fulfills numerous essential functions by forming a physical, chemical, and biological protective boundary with the external environment, besides acting as a sensory and endocrine organ. It is composed of the epidermis, a constantly renewed stratified epithelium, and the dermis, which harbors skin appendages such as hair follicles, sebaceous glands (SG), and sweat glands within a fibroblast-rich stroma (Fig. 1A). SGs are hair follicle-associated exocrine glands, whose lipid-rich secretion (sebum) primarily lubricates and protects the skin and hairs [1, 2]. In addition to the hair follicle-associated SGs, specialized SGs at specific body sites exert important functions. A prime example is found in the eyelids, where Meibomian glands secret meibum, the lipid fraction of the tear film that is essential for corneal health [3]. Sebum lipids show great variability across different mammals [4], very likely reflecting functional differences. In fury animals, sebum composition adapts to specific needs such as waterproofing, while in humans the sebum’s functions remain controversial. Human sebum has an unusual lipid composition, containing triglycerides, diglycerides, and free fatty acids (57%), wax esters (26%), squalene (12%), and cholesterol (2%) [1]. Wax esters and squalene are typical for sebum and are normally not found elsewhere in the body, and sebum fatty acids show uncommon saturation and branching patterns [5]. Deregulated sebum secretion is a key pathogenic factor in acne, the most common skin disease worldwide and a major burden during adolescence [6, 7] but it is also involved in several other debilitating skin diseases such as atopic dermatitis or psoriasis [8].
Fig. 1
Structure of of the pilosebaceous unit and cellular architecture of the sebaceous gland. A Schematic drawing of a pilosebaceous unit in sagittal section. B Higher magnification image of the sebaceous gland. The rectangle is enlarged in C to demonstrate the peripheral (SEB-B), differentiation (SEB-1–3) and necrosis (SEB-4) zones
However, instead of being simple fat-secreting glands, SGs are adaptable structures that respond to local and systemic stimuli, and constitute an appealing paradigm for research in topics beyond skin biology including stem cell function, adhesion and metabolism [9], the regulation of organ size [10], lipid metabolism [11], host-microbe interaction [12], whole body lipid and energetic metabolism [13] and tumorigenesis [14]. SG cells are interconnected at their lateral and basal surfaces by a variety of junctions that undergo dynamic rearrangements as cells differentiate. In spite of the increasing interest in studying SG pathophysiology, sebocyte cell adhesion processes and the underlying junctional complexes have been only superficially examined, and mostly as a by-product of studies focussing in other skin structures. Thus, systematic studies focussing on the SG are not available despite the general importance of cell–cell and cell–matrix adhesion in biological processes as embryonic development [15], and in particular in the epidermis [16]. In the latter tissue, for instance, corneodesmosomes formed by desmosomal components crosslinked to the cornified envelope bind corneocytes together and are essential for barrier function [16,17,18]. Importantly, changes in the expression of corneodesmosin, a major component of corneodesmosomes, have been reported in several skin diseases including ichtyoses, psoriasis, atopic dermatitis and benign and maligna skin tumors [19,20,21]. In addition, severe structural changes in SGs were reported after loss of adhesion-relevant proteins in genetically modified mice [22] (see also Table 1), strongly indicating an essential role for such components in the SG. This essay will highlight studies addressing structural and functional aspects of cell–cell and cell–matrix adhesion in SGs.
Table 1 Changes in sebaceous gland morphology and function in genetically modified mice with altered expression of junctional proteinsSebaceous gland development, morphology and functionSGs are usually found in the skin attached to hair follicles (the pilosebaceous unit), and their secretion coats and moistures the skin and hairs. SG development is intimately connected to hair follicle morphogenesis [36]. Markers of future stem cell compartments, including SRY-box transcription factor 9 (SOX9) and leucine-rich repeats and immunoglobulin-like domain protein 1 (LRIG1), are already expressed early stages of hair follicle formation [37, 38]. As hair follicle morphogenesis and lineage specification progresses, SOX9+ keratinocytes migrate towards the future bulge, while LRIG1+ cells locate at the upper part of the developing follicle, the future junctional zone [37]. The first sebocytes are generated in the junctional zone by asymmetric cell fate decision of proliferative Lrig1+ stem cells [39, 40]. Mature sebocytes are continuously replaced by dividing progenitor cells at the gland’s periphery, but the molecular mechanisms underlying SG cellular turnover remain to be elucidated. While LRIG1+ keratinocytes of the junctional zone of the hair follicle are the best-characterized stem cell population linked to SG renewal, sebocytes can also be derived from additional skin stem cell pools, including leucine-rich repeat-containing G-protein-coupled receptor 6+ progenitors and keratin 15+ stem cells of the upper bulge/isthmus region [37, 38].
Adult SGs (Fig. 1A) usually consist of a single lobule (acinus) or multiple lobules that open into the so-called sebaceous duct, which in turn disembogues into the pilary canal (free sebaceous glands open directly onto the skin’s surface). The glands are enclosed by a connective tissue capsule, formed by a basal lamina and its surrounding extracellular matrix (ECM), that gives rise to a system of trabeculae that separates the various acini and provide structural support [2, 41]. The surrounding stroma is composed of collagenous fibers and fibroblasts, and includes blood vessels and nerves.
Acinar cells show a progressive differentiation (Fig. 1B), which begins at the most peripheral layer and ends in its middle, when lipid-filled sebocytes disrupt and release their cellular contents in a process termed holocrine secretion [2, 42]. Although it is a continuous process, holocrine secretion has been traditionally divided in discrete stages based on the cellular morphology, most commonly in peripheral, maturation, and necrosis zones [41, 43]. More recent studies based on transcriptome data [44, 45] allowed establishing stages with higher resolution (Fig. 1C), including SEB-B for the flat, peripheral and in part mitotically active cells in contact with the basal lamina, SEB-1 and SEB-2 for initial and advanced stage of cells lost contact to the basal lamina and show massive increase in cell volume due to cytoplasmic lipid droplet accumulation, and SEB-3, the most advanced maturation stage, which embraces cells undergoing disruption. A fifth stage (SEB-4) comprising even more advanced cells, but whose transcripts are not assessable anymore, is the immediate source of lipids and cellular debris that reaches the skin surface. This process is guided by fine-tuned changes in the expression of selected genes [2]. From the peripheral layer to the gland’s middle, the transcriptional differentiation programme is initially characterized by proliferation and oxidative phosphorylation, followed by lipid metabolism and finally apoptosis and lysis [45]. One of the best characterized regulators of lipid synthesis in adult sebocytes is stearoyl-CoA desaturase 1 (Scd1), a rate-limiting enzyme in the synthesis of monounsaturated fatty acids, and naturally ocurring or experimentally induced mutations of the Scd1 gene result in absent or strongly hyoplastic SG and meibomian glands [46].
General aspects of cell adhesion structuresCell–cell adhesionCell–cell adhesion refers to the process of cells sticking to another cell. Adhesion between adjacent cells is mediated by a variety of structures, including tight junctions (TJ), adherens junctions (AJ), desmosomes, and gap junctions (GJ) (Fig. 2).
Fig. 2
Overview of cell–cell and cell–matrix adhesion structures
DesmosomesDesmosomes are intercellular junctions that anchor intermediate filaments to the plasma membrane [47, 48]. They are found primarily in tissues that are subjected to high mechanical stress, such as the myocardium and epidermis, but are also present in the gastrointestinal tract mucosa, the bladder, the meninges, and the dendritic reticulum of lymphatic follicles. The intercellular junctions are composed by proteins from the cadherin family, desmogleins and desmocollins, while on the cytoplasmic side, attachment plaques, including desmoplakins and plakoglobins, secure the intermediate filaments. In addition to its function in cell–cell adhesion, desmosomes perform a variety of roles in processes including cell signaling, differentiation, and tumor formation [49].
Adherens junctionsAJ-based extracellular adhesive contacts between cells serve to maintain tissue cohesion, sense and respond to tensile forces at the contact interface, establish cell polarity, and form intracellular links to cytoskeletal elements [50,51,52]. AJs are capable of localizing proteins to subcellular compartments, which allows for the modulation of signaling pathways. AJs comprise three main components: transmembrane cadherins, armadillo family members, and cytoskeletal adaptor proteins. This core cadherin-catenin complex binds to the actomyosin cytoskeleton and signaling proteins, thereby influencing the overall mechanobiology of cells. In particular, it plays a key role in the integrations of cell proliferation, fate, and positioning to establish the boundary between basal and suprabasal layers of epithelial tissues [53].
Tight junctionsTJs, also designated zonulae occludens, seal the paracellular space, thereby creating a permeability barrier that is nearly leak-proof [54]. TJs have two main functions: the fence and gate functions [55]. In terms of the fence function, TJ serves to establish membrane polarity by separating the plasma membrane into apical and basolateral domains, thereby creating an asymmetry regarding the composition of cytosol and plasma membrane proteins and lipids. With regard to the gate function, TJ establish a paracellular diffusion barrier between sealed cells and regulates the passage of solutes and ion selectivity. Furthermore, TJ regulates the organization of cytoskeletal proteins, controlling the actomyosin contractility and distributing cytoskeletal-generated tensional forces [50]. TJ comprises three main transmembrane proteins: junctional adhesion molecules (JAM), claudin, and occludin. Scaffolding proteins, such as zonula occludens proteins (ZO-1, ZO-2, ZO-3), and cingulin associate occludin, claudin, and JAM in tight junctional strands, promoting polymerization.
Gap junctionsGJs are essential membrane proteins that regulate the cellular response of heterogeneous cells. They function as intercellular channels of communication, facilitating the transport of small molecules such as amino acids, sugars, intracellular messengers and ions between cells [56]. The fundamental structural units of GJ are connexins. Six connexin proteins are arranged around a pore in a hexagonal configuration, forming a structure called a connexon [57]. Individual connexons (hemichannels) containing a single connexin type are referred to as homomeric, while those comprising different connexins are classified as heteromeric.
Cell–matrix adhesionCell–matrix adhesion refers to the connection of cells with the extracellular matrix (ECM). This interaction regulates numerous processes beyond cell adhesion, including migration, signaling during morphogenesis, tissue homeostasis, wound healing, and tumorigenesis [58]. Though extremely heterogeneous in its composition, animal ECM largely falls into two categories: the basement membrane (BM) and in interstitial matrix. The BM, a dense thin layer of ECM that marks the boundary of many tissues, consists of laminin and collagen IV networks and crosslinking molecules such as nidogen and perlecan, besides other proteins as fibronectin, tenascin C, fibrillin, agrin, and collagens XV and XVIII. Besides anchoring cellular sheets, the BM has important roles in guiding tissue morphogenesis [59]. In the interstitial matrix, collagens and other proteins such as fibronectin, elastin, laminin, and tenascin, build a characteristic, fibrous network, while glycosaminoglycans, proteoglycans and water contribute to their interstitial spaces [59, 60]. A simplified view of the contribution of these different ECM components to the microenvironmental mechanical properties is that glycosaminoglycans and proteins are responsible for the ECM compressive and tensile strength, respectively [59].
Cell adhesion to ECM substrates is largely mediated by integrin adhesion complexes (IACs), clusters of integrins with associated signaling, scaffolding and cytoskeletal proteins [61]. Integrins are linked via adapter proteins to the actin filaments or intermediate filaments of the own cell, and connect to the BM by binding to a variety of ligands including fibronectin, vitronectin, collagen and laminin. IACs are very heterogeneous and have been classified based on criteria as size, composition, lifetime, cellular distribution and function, and include focal adhesions (FAs), focal complexes, fibrillar adhesions, reticular adhesions, invadosomes, and hemidesmosomes (HDs), among others [61]. The most extensively studied and best characterized cell–matrix adhesions structures are FAs and HDs (Fig. 2). A crosstalk between Fas and HDs integrates mechanotransduction at cell–cell and cell–ECM adhesions and regulates several aspects of cellular behavior, including cell migration and tissue development [62].
Cell adhesion in sebaceous glandsNumerous cell–cell and cell–matrix adhesion structures have been described in SG cells via a variety of methods including electron microscopy, immunohistochemistry, and immunofluorescence. Recent single-cell transcriptomics studies confirmed the expression of a large number of adhesion-relevant transcripts in both mouse [63] and human [64] SGs. Figure 3A, based on a re-assessment of these data focusing on sebocytes [65], indicates the number of adhesion-relevant transcripts that are expressed at high and low levels in sebocytes compared to other skin cell types. Assessment of another recent dataset based on the spatial transcriptomics analysis of normal and diseased human skin [66, 67] also indicates substantial shifts in the abundance of adhesion-relevant transcripts in psoriasis and atopic dermatitis lesions compared to healthy skin (Fig. 3B).
Fig. 3
Comparison of gene expression associated to the gene ontology terms on cell–cell interaction and cell–matrix interaction . A Number of adhesion molecules with weak (yellow) or strong (green) expression (legend: below) in mouse (left) and human (right) sebaceous glands according to single-cell transcription studies. This information is retrieved from existing data published together with our previous study [65]. A gene is strong expressed, if its mean expression over all sebaceous gland samples annotated in the original experimental studies [63, 64] is greater or equal compared to the mean expression of any sample. Gene ontology identifier, terms and corresponding gene sets have been annotated within our previous study [65]. B Difference in base-10 logarithmic mean gene expression for matched-pair lesion and non-lesion samples [66]: Atopic dermatitis (left, 4 patients) and psoriasis (right, 3 patients). Each dot represents one gen from one matched-pair, thus redundant gene annotation occurs. Genes with a difference in mean expression greater than 1 between lesion (red squares) and non-lesion (blue triangles) are highlighted (legend: below) for atopic dermatitis and psoriasis. For psoriasis no non-lesion sample with this difference occurs
In the next sections, we will highlight structural and functional roles reported for adhesive structures in SGs.
Cell–cell adhesion in sebaceous glandsDesmosomesVarious electron microscope-based studies in humans and other species [43, 68,69,70,71] reported the presence of desmosomes in SG cells. These reports indicate that desmosomes are quite numerous in basal cells, less frequent in mature cells, and rare or absent in fully differentiated cells (Fig. 4). Several desmosomal components were detected in SGs by immunohistochemistry, including desmogleins, desmoplakin, plakoglobin, and desmocollin. Among these, desmoglein 1 was reported to be found in excretory duct epithelial cells, basal cells, differentiating cells, and mature cells of human SG [72]. Notably, while desmoglein-1-alpha (DSG1A) was completely absent in the SGs of adult mouse ears, DSG1B was expressed in the outer epithelial lining and DSG1G was expressed throughout the gland, including the more differentiated cell layers [73]. Desmoplakin, another major desmosomal protein, stained positive at the cell–cell contacts along the entire SG differentiation axis [26]. Additionally, desmoplakin, desmocollin 3, and placoglobin were found in basal cells, differentiated cells, and mature cells of sebaceous glands [26]. Mutations in genes encoding desmos
留言 (0)